

doi: 10.1073/pnas.0135345100.
Epub 2002 Dec 16.
Bidirectional synaptic plasticity in the cerebellum-like mammalian dorsal cochlear nucleus
Affiliations
- PMID: 12486245
- PMCID: PMC140946
- DOI: 10.1073/pnas.0135345100
Item in Clipboard
Bidirectional synaptic plasticity in the cerebellum-like mammalian dorsal cochlear nucleus
Proc Natl Acad Sci U S A.
.
Abstract
The dorsal cochlear nucleus integrates acoustic with multimodal sensory inputs from widespread areas of the brain. Multimodal inputs are brought to spiny dendrites of fusiform and cartwheel cells in the molecular layer by parallel fibers through synapses that are subject to long-term potentiation and long-term depression. Acoustic cues are brought to smooth dendrites of fusiform cells in the deep layer by auditory nerve fibers through synapses that do not show plasticity. Plasticity requires Ca(2+)-induced Ca(2+) release; its sensitivity to antagonists of N-methyl-d-aspartate and metabotropic glutamate receptors differs in fusiform and cartwheel cells.
Figures
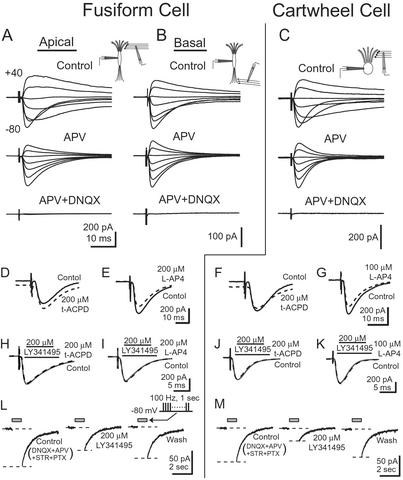
Glutamatergic receptors of the AMPA, NMDA, and mGluR subtypes contribute to synaptic responses in fusiform and cartwheel cells. (A) In a fusiform cell, EPSCs were evoked by stimulating the molecular layer while the holding potential of the cell was varied between −80 and +40 mV in 20-mV steps. A slow component that was prominent at depolarizing holding potentials was blocked by 50 μM APV, indicating that it was mediated through NMDA receptors. The remaining fast component was blocked by 20 μM DNQX, showing that it was mediated through AMPA receptors. (B) In a different fusiform cell, EPSCs were evoked through the deep layer. EPSCs contained currents through both NMDA and AMPA receptors. (C) In a cartwheel cell, EPSCs evoked by shocks to the molecular layer had a relatively large current through NMDA receptors. (D) In a fusiform cell, 200 μM t-ACPD shifted the holding current at −80 mV inwardly but did not change the amplitude of the EPSC evoked by a shock to parallel fibers. (E) Two hundred micromolar L-AP4 decreased the amplitude of EPSCs evoked by stimulating parallel fibers but did not shift the holding current. (F) In a cartwheel cell, 200 μM t-ACPD shifted the holding current inwardly and decreased the amplitude of the EPSCs. (G) In another cartwheel cell, 100 μM L-AP4 decreased the amplitude of EPSCs without shifting the holding current. (H–K) Maximal responses to either t-ACPD or L-AP4 were blocked by LY341495. Solid gray line shows response to parallel fiber stimulation in the presence of 200 μM LY341495. Dashed black line shows response in the additional presence of agonist. (L and M) Train of shocks to parallel fibers, 100 Hz, 1 sec [gray bar (Inset), with stimulus artifacts removed], reveals a current mediated by mGluR. A slow inward current after the shocks was evoked in the presence of 20 μM DNQX and 100 μM APV at −80 mV. The inward current was partially but reversibly blocked by 200 μM LY341495.
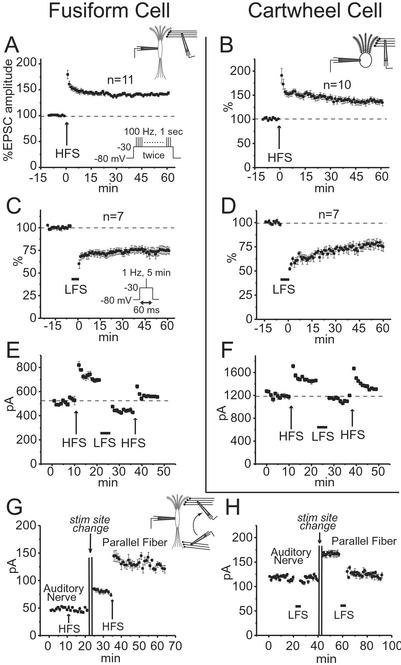
Bidirectional plasticity was observed in responses to shocks of parallel fibers in fusiform and cartwheel cells but not in responses to shocks of auditory nerve fibers. (A and B) LTP was evoked in fusiform and cartwheel cells by HFS. HFS comprised two repetitions with a 20-sec interval of depolarizing the postsynaptic cell from −80 to −30 mV for 1 sec while stimulating parallel fibers for 100 Hz (A Inset). Plots show mean, normalized EPSC amplitudes from all cells in which recordings were made for >60 min after HFS. (C and D) LTD was evoked in fusiform and cartwheel cells by LFS. LFS comprised stimulating parallel fibers at 1 Hz for 5 min, each shock being presented during a 60-ms depolarization of the postsynaptic cell from −80 to −30 mV (C Inset). Plots show mean normalized EPSC amplitudes from all cells in which recordings were made for >60 min after LFS. (E and F) In a fusiform and a cartwheel cell, potentiation and depression could be reversed. (G) In a fusiform cell, LTP could not be induced by stimulation in the deep layer, but could subsequently be induced by moving the stimulating electrode to the molecular layer. (H) In a fusiform cell, LTD could not be induced with stimulation in the deep layer, but could subsequently be induced by moving the stimulating pipette to the molecular layer.
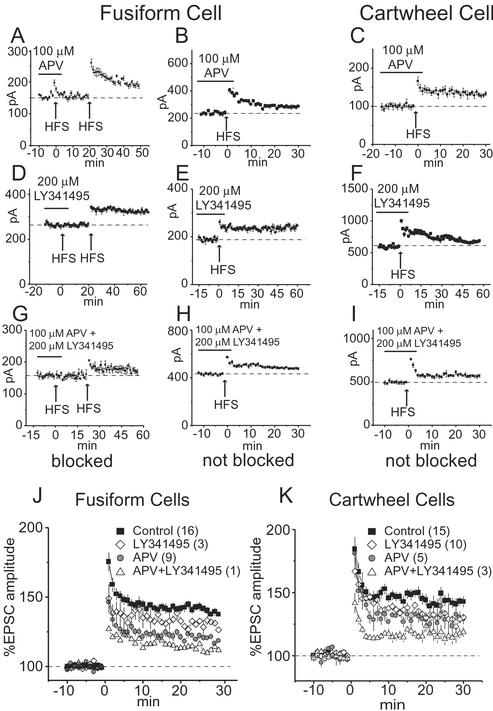
NMDA and mGluRs contribute to the induction of LTP in fusiform and cartwheel cells. (A) In a fusiform cell, LTP was blocked by 100 μM APV but could subsequently be induced after APV was washed out of the bath. (B) In another fusiform cell, APV did not block the induction of LTP. (C) In a cartwheel cell, LTP was not blocked by APV. (D) In a fusiform cell, 200 μM LY341495 reversibly blocked LTP. (E) In another fusiform cell, LY341495 did not block LTP. (F) In a cartwheel cell, LY341495 failed to block LTP. (G and H) Simultaneous application of APV and LY341495 blocked LTP in one but not another fusiform cell. (I) The combination of APV and LY341495 did not block LTP in any cartwheel cell. (J and K) Plots show the mean normalized amplitudes of EPSCs for the control and the presence of LY341495 and/or APV during the conditioning stimuli. The number of cells whose responses were averaged is given in parentheses.
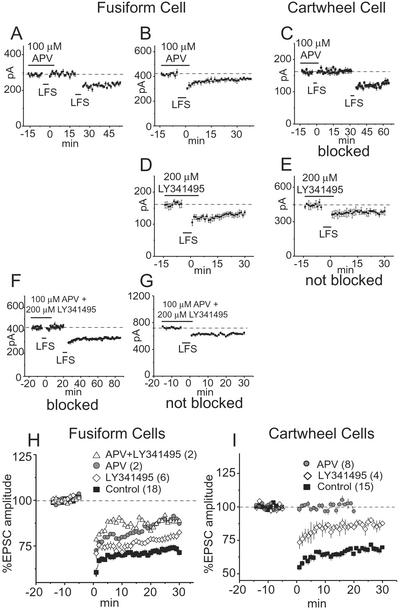
NMDA and mGluRs contribute to the induction of LTD in fusiform and cartwheel cells. (A) In a fusiform cell, the induction of LTD by LFS was blocked by 100 μM APV but could be evoked after APV was washed out of the bath. (B) In another fusiform cell, LTD was not blocked by APV. (C) In all cartwheel cells tested, LTD was reversibly blocked by APV. (D) In all fusiform cells tested, 200 μM LY341495 failed to block LTD. (E) In all cartwheel cells, LY341495 failed to block LTD. (F and G) The simultaneous application of APV and LY341495 blocked LTD in some fusiform cells. (H and I) Averaged normalized EPSCs were compared in populations of cells. Each plot shows the mean normalized amplitudes of EPSCs in control conditions and in the presence of LY341495 and/or APV. The number of cells whose responses were averaged is given in parentheses.
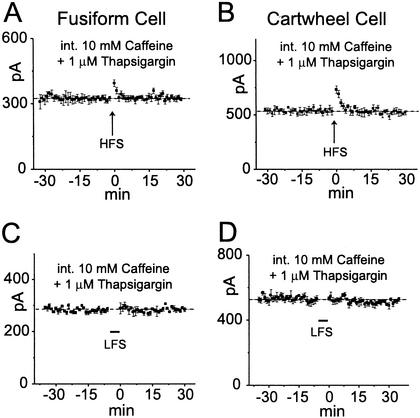
Synaptic plasticity in fusiform and cartwheel cells involves Ca2+-induced Ca2+ release. The recording pipettes contained 10 mM caffeine to release Ca2+ from intracellular stores and 1 μM thapsigargin to block its re-uptake. Conditioning stimuli were given after a prolonged control period (30 min) to allow the drugs to diffuse into distal dendrites.
Similar articles
-
Cell-specific, spike timing-dependent plasticities in the dorsal cochlear nucleus.Nat Neurosci. 2004 Jul;7(7):719-25. doi: 10.1038/nn1272. Epub 2004 Jun 20. Nat Neurosci. 2004. PMID: 15208632
-
Plasticity of synaptic GluN receptors is required for the Src-dependent induction of long-term potentiation at CA3-CA1 synapses.Hippocampus. 2011 Oct;21(10):1053-61. doi: 10.1002/hipo.20818. Epub 2010 Jun 2. Hippocampus. 2011. PMID: 20865743
-
Induction of homosynaptic long-term depression at spinal synapses of sensory a delta-fibers requires activation of metabotropic glutamate receptors.Neuroscience. 2000;98(1):141-8. doi: 10.1016/s0306-4522(00)00080-4. Neuroscience. 2000. PMID: 10858620
-
Mechanisms of synaptic plasticity in the dorsal cochlear nucleus: plasticity-induced changes that could underlie tinnitus.Am J Audiol. 2008 Dec;17(2):S170-5. doi: 10.1044/1059-0889(2008/07-0030). Epub 2008 Oct 31. Am J Audiol. 2008. PMID: 18978197 Free PMC article. Review.
-
What's a cerebellar circuit doing in the auditory system?Trends Neurosci. 2004 Feb;27(2):104-10. doi: 10.1016/j.tins.2003.12.001. Trends Neurosci. 2004. PMID: 15102490 Review.
Cited by
-
Coactivation of pre- and postsynaptic signaling mechanisms determines cell-specific spike-timing-dependent plasticity.Neuron. 2007 Apr 19;54(2):291-301. doi: 10.1016/j.neuron.2007.03.026. Neuron. 2007. PMID: 17442249 Free PMC article.
-
Dopaminergic modulation of axon initial segment calcium channels regulates action potential initiation.Neuron. 2010 Nov 4;68(3):500-11. doi: 10.1016/j.neuron.2010.09.026. Neuron. 2010. PMID: 21040850 Free PMC article.
-
Activation of synaptic group II metabotropic glutamate receptors induces long-term depression at GABAergic synapses in CNS neurons.J Neurosci. 2013 Oct 2;33(40):15964-77. doi: 10.1523/JNEUROSCI.0202-13.2013. J Neurosci. 2013. PMID: 24089501 Free PMC article.
-
Anatomy and Physiology of Metabotropic Glutamate Receptors in Mammalian and Avian Auditory System.HSOA Trends Anat Physiol. 2018;1:001. doi: 10.24966/TAP-7752/100001. Epub 2018 Feb 9. HSOA Trends Anat Physiol. 2018. PMID: 30854519 Free PMC article.
-
Plasticity of somatosensory inputs to the cochlear nucleus--implications for tinnitus.Hear Res. 2011 Nov;281(1-2):38-46. doi: 10.1016/j.heares.2011.05.001. Epub 2011 May 18. Hear Res. 2011. PMID: 21620940 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
