

The yeast DHHC cysteine-rich domain protein Akr1p is a palmitoyl transferase
- PMID: 12370247
- PMCID: PMC2173492
- DOI: 10.1083/jcb.200206120
The yeast DHHC cysteine-rich domain protein Akr1p is a palmitoyl transferase
Abstract
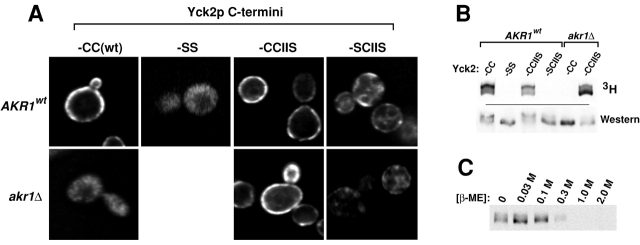
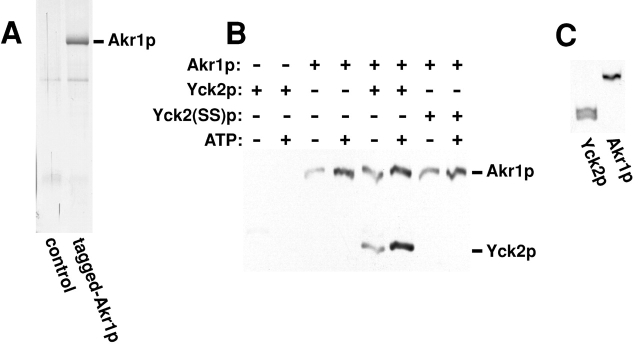
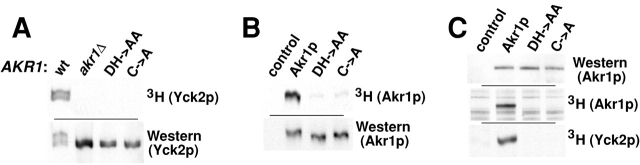
Protein palmitoylation has been long appreciated for its role in tethering proteins to membranes, yet the enzymes responsible for this modification have eluded identification. Here, experiments in vivo and in vitro demonstrate that Akr1p, a polytopic membrane protein containing a DHHC cysteine-rich domain (CRD), is a palmitoyl transferase (PTase). In vivo, we find that the casein kinase Yck2p is palmitoylated and that Akr1p function is required for this modification. Akr1p, purified to near homogeneity from yeast membranes, catalyzes Yck2p palmitoylation in vitro, indicating that Akr1p is itself a PTase. Palmitoylation is stimulated by added ATP. Furthermore, during the reaction, Akr1p is itself palmitoylated, suggesting a role for a palmitoyl-Akr1p intermediate in the overall reaction mechanism. Mutations introduced into the Akr1p DHHC-CRD eliminate both the trans- and autopalmitoylation activities, indicating a central participation of this conserved sequence in the enzymatic reaction. Finally, our results indicate that palmitoylation within the yeast cell is controlled by multiple PTase specificities. The conserved DHHC-CRD sequence, we propose, is the signature feature of an evolutionarily widespread PTase family.
Figures
Similar articles
-
Transmembrane topology of the protein palmitoyl transferase Akr1.J Biol Chem. 2005 Mar 18;280(11):10156-63. doi: 10.1074/jbc.M411946200. Epub 2005 Jan 4. J Biol Chem. 2005. PMID: 15632165
-
Akr1p-dependent palmitoylation of Yck2p yeast casein kinase 1 is necessary and sufficient for plasma membrane targeting.J Biol Chem. 2004 Jun 25;279(26):27138-47. doi: 10.1074/jbc.M403071200. Epub 2004 Apr 22. J Biol Chem. 2004. PMID: 15105419
-
Model organisms lead the way to protein palmitoyltransferases.J Cell Sci. 2004 Feb 1;117(Pt 4):521-6. doi: 10.1242/jcs.00989. J Cell Sci. 2004. PMID: 14730009
-
Protein Palmitoylation by DHHC Protein Family.In: Kittler JT, Moss SJ, editors. The Dynamic Synapse: Molecular Methods in Ionotropic Receptor Biology. Boca Raton (FL): CRC Press/Taylor & Francis; 2006. Chapter 5. In: Kittler JT, Moss SJ, editors. The Dynamic Synapse: Molecular Methods in Ionotropic Receptor Biology. Boca Raton (FL): CRC Press/Taylor & Francis; 2006. Chapter 5. PMID: 21204476 Free Books & Documents. Review.
-
Systematic screening for palmitoyl transferase activity of the DHHC protein family in mammalian cells.Methods. 2006 Oct;40(2):177-82. doi: 10.1016/j.ymeth.2006.05.015. Methods. 2006. PMID: 17012030 Review.
Cited by
-
Profiling targets of the irreversible palmitoylation inhibitor 2-bromopalmitate.ACS Chem Biol. 2013 Sep 20;8(9):1912-7. doi: 10.1021/cb400380s. Epub 2013 Jul 25. ACS Chem Biol. 2013. PMID: 23844586 Free PMC article.
-
Cu/Zn superoxide dismutase and the proton ATPase Pma1p of Saccharomyces cerevisiae.Biochem Biophys Res Commun. 2015 Jul 3;462(3):251-6. doi: 10.1016/j.bbrc.2015.04.127. Epub 2015 May 6. Biochem Biophys Res Commun. 2015. PMID: 25956063 Free PMC article.
-
In silico screening for palmitoyl substrates reveals a role for DHHC1/3/10 (zDHHC1/3/11)-mediated neurochondrin palmitoylation in its targeting to Rab5-positive endosomes.J Biol Chem. 2013 Jul 5;288(27):19816-29. doi: 10.1074/jbc.M112.431676. Epub 2013 May 16. J Biol Chem. 2013. PMID: 23687301 Free PMC article.
-
spe-10 encodes a DHHC-CRD zinc-finger membrane protein required for endoplasmic reticulum/Golgi membrane morphogenesis during Caenorhabditis elegans spermatogenesis.Genetics. 2006 Jan;172(1):145-58. doi: 10.1534/genetics.105.047340. Epub 2005 Sep 2. Genetics. 2006. PMID: 16143610 Free PMC article.
-
The vacuolar DHHC-CRD protein Pfa3p is a protein acyltransferase for Vac8p.J Cell Biol. 2005 Sep 26;170(7):1091-9. doi: 10.1083/jcb.200507048. J Cell Biol. 2005. PMID: 16186255 Free PMC article.
References
-
- Berthiaume, L., and M.D. Resh. 1995. Biochemical characterization of a palmitoyl acyltransferase activity that palmitoylates myristoylated proteins. J. Biol. Chem. 270:22399–22405. - PubMed
-
- Bizzozero, O.A. 1995. Chemical analysis of acylation sites and species. Methods Enzymol. 250:361–379. - PubMed
-
- Brown, D.A., and E. London. 2000. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J. Biol. Chem. 275:17221–17224. - PubMed
-
- Campbell, S.M., S.M. Crowe, and J. Mak. 2001. Lipid rafts and HIV-1: from viral entry to assembly of progeny virions. J. Clin. Virol. 22:217–227. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
