

Distinct domains of the GATA-1 cofactor FOG-1 differentially influence erythroid versus megakaryocytic maturation
- PMID: 12024038
- PMCID: PMC133877
- DOI: 10.1128/MCB.22.12.4268-4279.2002
Distinct domains of the GATA-1 cofactor FOG-1 differentially influence erythroid versus megakaryocytic maturation
Abstract
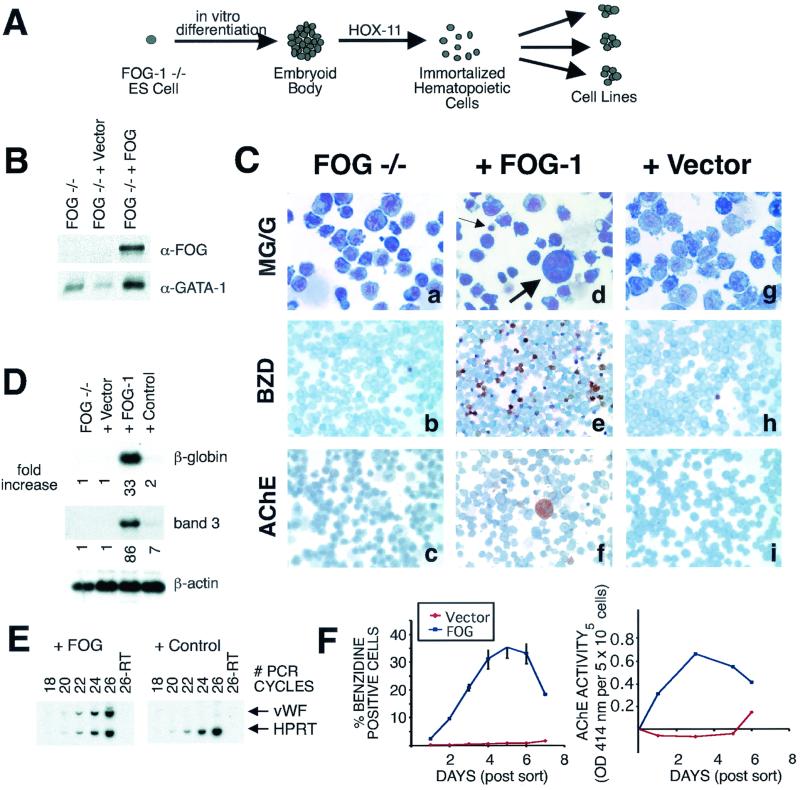
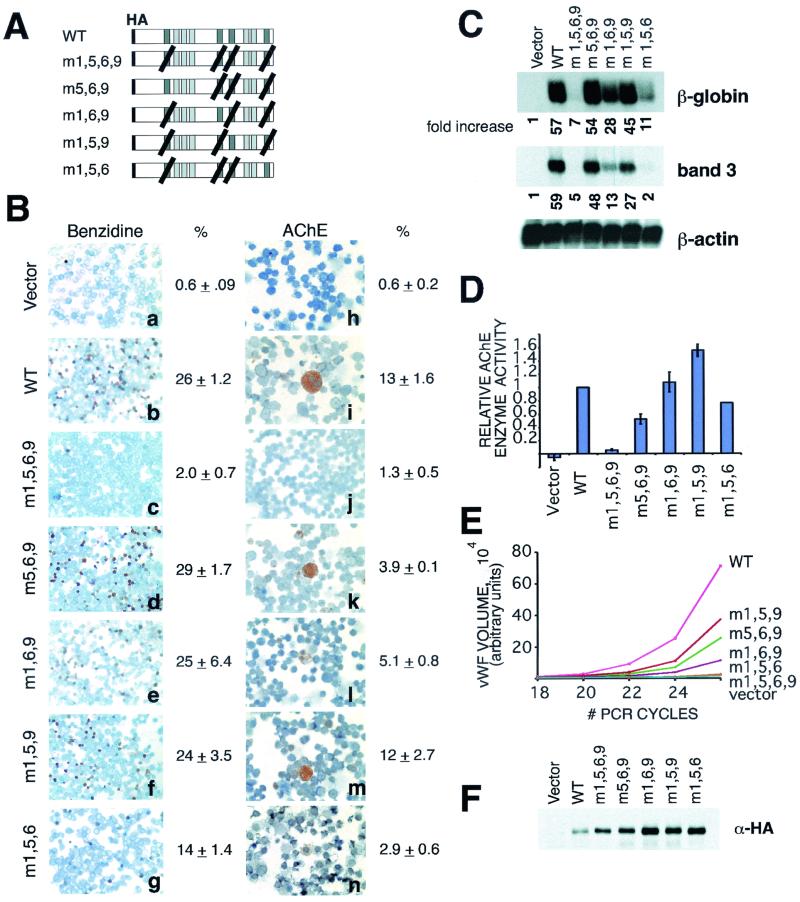
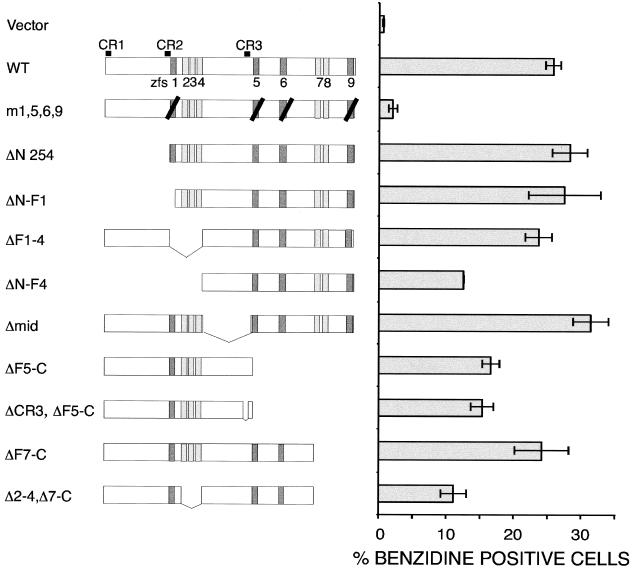
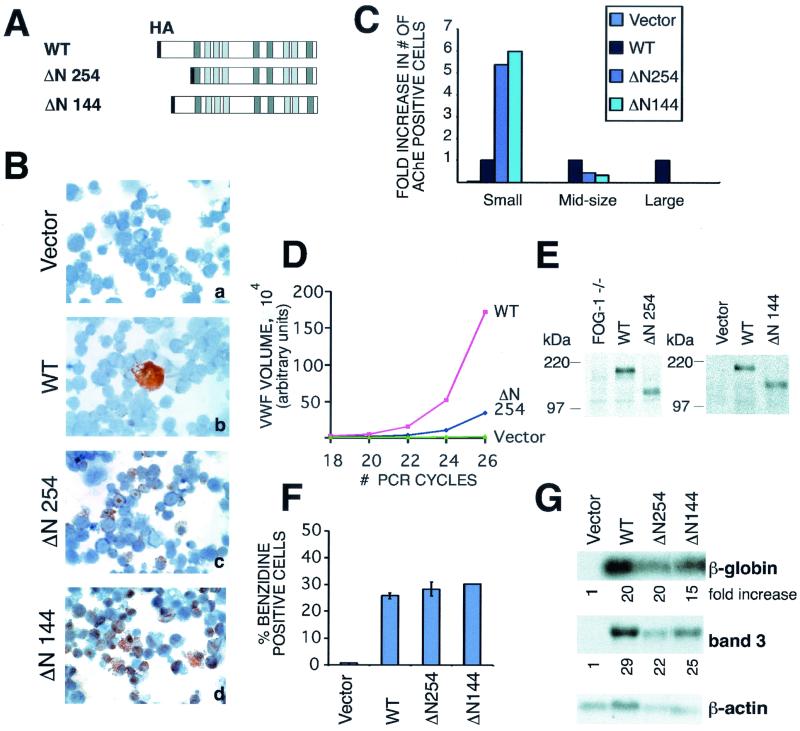
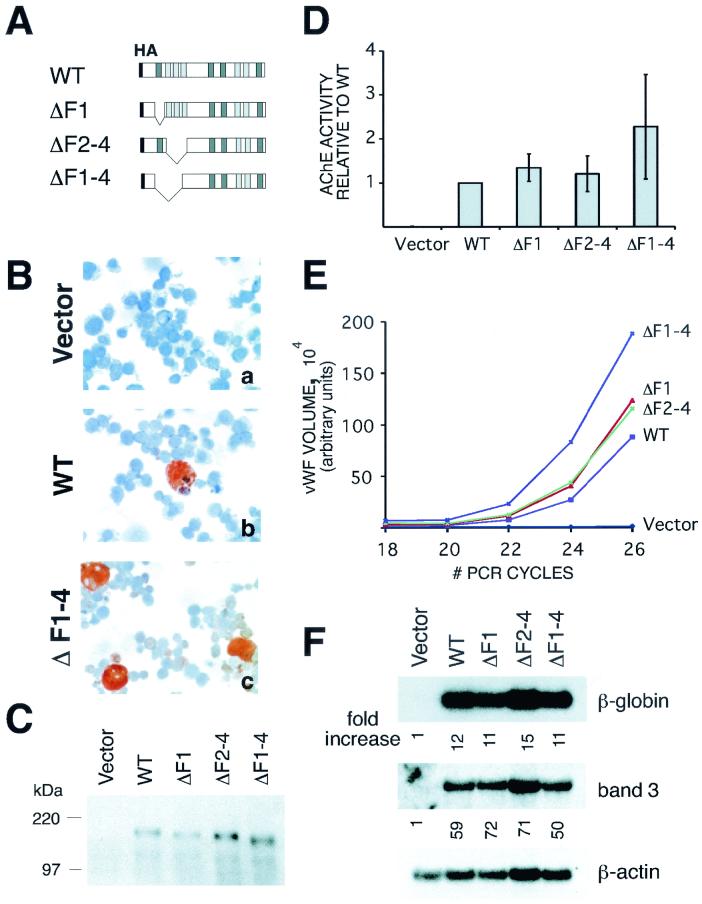
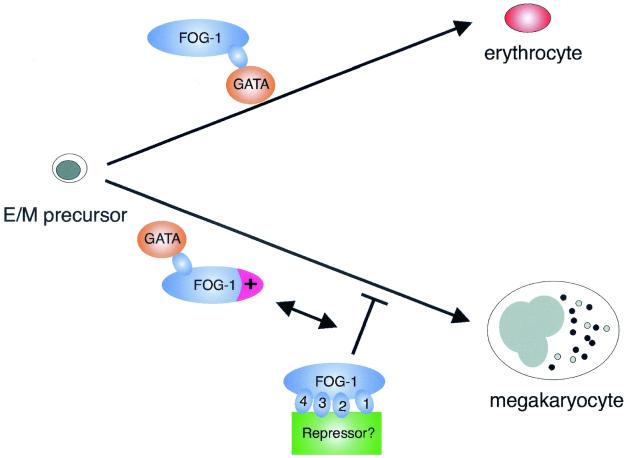
FOG family zinc finger proteins play essential roles in development through physical interaction with GATA factors. FOG-1, like its interacting partner GATA-1, is required for normal differentiation of erythroid and megakaryocytic cells. Here, we have developed a functional assay for FOG-1 based on its ability to rescue erythroid and megakaryocytic maturation of a genetically engineered FOG-1(-/-) cell line. We demonstrate that interaction through only one of FOG-1's four GATA-binding zinc fingers is sufficient for rescue, providing evidence against a model in which FOG-1 acts to bridge multiple GATA-binding DNA elements. Importantly, we find that distinct regions of FOG-1 differentially influence erythroid versus megakaryocyte maturation. As such, we propose that FOG-1 may modulate the fate of a bipotential erythroid/megakaryocytic precursor cell.
Figures
Similar articles
-
FOG, a multitype zinc finger protein, acts as a cofactor for transcription factor GATA-1 in erythroid and megakaryocytic differentiation.Cell. 1997 Jul 11;90(1):109-19. doi: 10.1016/s0092-8674(00)80318-9. Cell. 1997. PMID: 9230307
-
Use of altered specificity mutants to probe a specific protein-protein interaction in differentiation: the GATA-1:FOG complex.Mol Cell. 1999 Feb;3(2):219-28. doi: 10.1016/s1097-2765(00)80312-3. Mol Cell. 1999. PMID: 10078204
-
Control of megakaryocyte-specific gene expression by GATA-1 and FOG-1: role of Ets transcription factors.EMBO J. 2002 Oct 1;21(19):5225-34. doi: 10.1093/emboj/cdf527. EMBO J. 2002. PMID: 12356738 Free PMC article.
-
Coregulation of GATA factors by the Friend of GATA (FOG) family of multitype zinc finger proteins.Semin Cell Dev Biol. 2005 Feb;16(1):117-28. doi: 10.1016/j.semcdb.2004.10.006. Epub 2004 Dec 15. Semin Cell Dev Biol. 2005. PMID: 15659346 Review.
-
Regulation of the transcription factor GATA-1 at the gene and protein level.Cell Mol Life Sci. 2001 Dec;58(14):2008-17. doi: 10.1007/PL00000833. Cell Mol Life Sci. 2001. PMID: 11814053 Free PMC article. Review.
Cited by
-
TLX1 (HOX11) immortalization of embryonic stem cell-derived and primary murine hematopoietic progenitors.Curr Protoc Stem Cell Biol. 2008 Dec;Chapter 1:Unit 1F.7. doi: 10.1002/9780470151808.sc01f07s7. Curr Protoc Stem Cell Biol. 2008. PMID: 19085976 Free PMC article.
-
Friend of GATA suppresses the GATA-induced transcription of hepcidin in hepatocytes through a GATA-regulatory element in the HAMP promoter.J Mol Endocrinol. 2011 Nov 21;47(3):299-313. doi: 10.1530/JME-11-0060. Print 2011 Dec. J Mol Endocrinol. 2011. PMID: 21971825 Free PMC article.
-
Combinatorial interactions of serpent, lozenge, and U-shaped regulate crystal cell lineage commitment during Drosophila hematopoiesis.Proc Natl Acad Sci U S A. 2003 Sep 30;100(20):11451-6. doi: 10.1073/pnas.1635050100. Epub 2003 Sep 22. Proc Natl Acad Sci U S A. 2003. PMID: 14504400 Free PMC article.
-
GATA switches as developmental drivers.J Biol Chem. 2010 Oct 8;285(41):31087-93. doi: 10.1074/jbc.R110.159079. Epub 2010 Jul 29. J Biol Chem. 2010. PMID: 20670937 Free PMC article. Review.
-
FOG-1-mediated recruitment of NuRD is required for cell lineage re-enforcement during haematopoiesis.EMBO J. 2010 Jan 20;29(2):457-68. doi: 10.1038/emboj.2009.368. Epub 2009 Dec 10. EMBO J. 2010. PMID: 20010697 Free PMC article.
References
-
- Bulger, M., and M. Groudine. 1999. Looping versus linking: toward a model for long-distance gene activation. Genes Dev. 13:2465-2477. - PubMed
-
- Crispino, J. D., M. Lodish, J. P. Mackay, and S. H. Orkin. 1999. Use of altered specificity mutants to probe a specific protein-protein interaction in differentiation: the GATA-1:FOG complex. Mol. Cell 3:219-228. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials