

SpSld3 is required for loading and maintenance of SpCdc45 on chromatin in DNA replication in fission yeast
- PMID: 12006645
- PMCID: PMC111119
- DOI: 10.1091/mbc.02-01-0006
SpSld3 is required for loading and maintenance of SpCdc45 on chromatin in DNA replication in fission yeast
Abstract
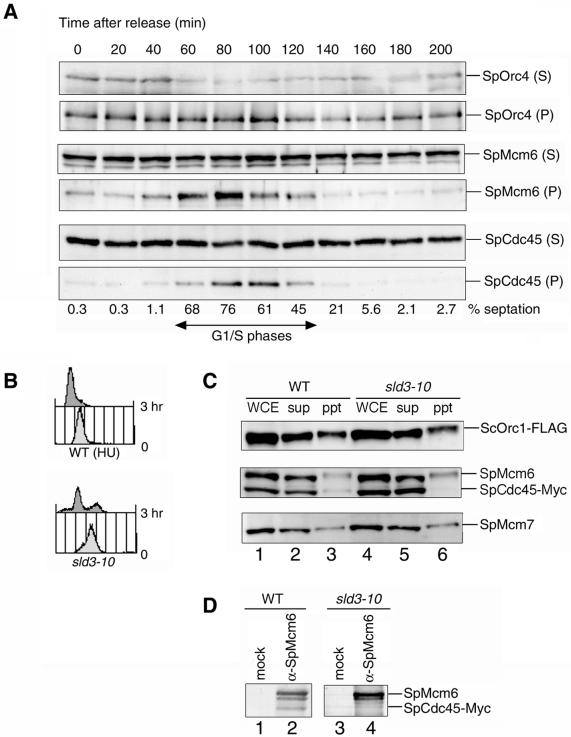
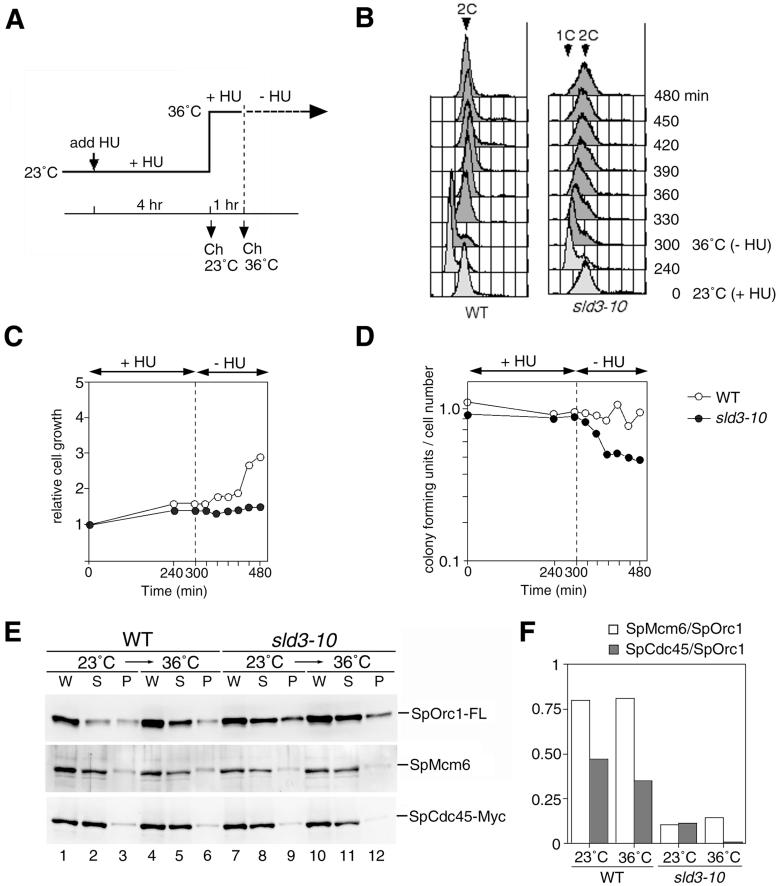
Initiation of DNA replication in eukaryotic cells is regulated through the ordered assembly of replication complexes at origins of replication. Association of Cdc45 with the origins is a crucial step in assembly of the replication machinery, hence can be considered a target for the regulation of origin activation. To examine the process required for SpCdc45 loading, we isolated fission yeast SpSld3, a counterpart of budding yeast Sld3 that interacts with Cdc45. SpSld3 associates with the replication origin during G1-S phases and this association depends on Dbf4-dependent (DDK) kinase activity. In the corresponding period, SpSld3 interacts with minichromosome maintenance (MCM) proteins and then with SpCdc45. A temperature-sensitive sld3-10 mutation suppressed by the multicopy of the sna41+ encoding SpCdc45 impairs loading of SpCdc45 onto chromatin. In addition, this mutation leads to dissociation of preloaded Cdc45 from chromatin in the hydroxyurea-arrested S phase, and DNA replication upon removal of hydroxyurea is retarded. Thus, we conclude that SpSld3 is required for stable association of Cdc45 with chromatin both in initiation and elongation of DNA replication. The DDK-dependent origin association suggests that SpSld3 is involved in temporal regulation of origin firing.
Figures
Similar articles
-
Sld3, which interacts with Cdc45 (Sld4), functions for chromosomal DNA replication in Saccharomyces cerevisiae.EMBO J. 2001 Apr 17;20(8):2097-107. doi: 10.1093/emboj/20.8.2097. EMBO J. 2001. PMID: 11296242 Free PMC article.
-
Ordered assembly of Sld3, GINS and Cdc45 is distinctly regulated by DDK and CDK for activation of replication origins.EMBO J. 2006 Oct 4;25(19):4663-74. doi: 10.1038/sj.emboj.7601347. Epub 2006 Sep 21. EMBO J. 2006. PMID: 16990792 Free PMC article.
-
Origin association of Sld3, Sld7, and Cdc45 proteins is a key step for determination of origin-firing timing.Curr Biol. 2011 Dec 20;21(24):2055-63. doi: 10.1016/j.cub.2011.11.038. Epub 2011 Dec 8. Curr Biol. 2011. PMID: 22169533
-
Origins and complexes: the initiation of DNA replication.J Exp Bot. 2001 Feb;52(355):193-202. J Exp Bot. 2001. PMID: 11283163 Review.
-
Cdc6 and DNA replication: limited to humble origins.Bioessays. 1996 Nov;18(11):859-62. doi: 10.1002/bies.950181103. Bioessays. 1996. PMID: 8939063 Review.
Cited by
-
Mcm10 plays an essential role in origin DNA unwinding after loading of the CMG components.EMBO J. 2012 May 2;31(9):2182-94. doi: 10.1038/emboj.2012.68. Epub 2012 Mar 20. EMBO J. 2012. PMID: 22433840 Free PMC article.
-
Phosphopeptide binding by Sld3 links Dbf4-dependent kinase to MCM replicative helicase activation.EMBO J. 2016 May 2;35(9):961-73. doi: 10.15252/embj.201593552. Epub 2016 Feb 24. EMBO J. 2016. PMID: 26912723 Free PMC article.
-
Switch on the engine: how the eukaryotic replicative helicase MCM2-7 becomes activated.Chromosoma. 2015 Mar;124(1):13-26. doi: 10.1007/s00412-014-0489-2. Epub 2014 Oct 12. Chromosoma. 2015. PMID: 25308420 Review.
-
Efficient initiation of DNA replication in eukaryotes requires Dpb11/TopBP1-GINS interaction.Mol Cell Biol. 2013 Jul;33(13):2614-22. doi: 10.1128/MCB.00431-13. Epub 2013 Apr 29. Mol Cell Biol. 2013. PMID: 23629628 Free PMC article.
-
Regulation of the initiation step of DNA replication by cyclin-dependent kinases.Chromosoma. 2010 Dec;119(6):565-74. doi: 10.1007/s00412-010-0291-8. Epub 2010 Aug 5. Chromosoma. 2010. PMID: 20686781 Review.
References
-
- Alfa C, Fantes P, Hyams J, Mcleod M, Warbrick E. Experiments with Fission Yeast, A Laboratory Course Manual. Cold Spring Harbor, NY.: Cold Spring Harbor Laboratory Press; 1993.
-
- Arata Y, Fujita M, Ohtani K, Kijima S, Kato JY. Cdk2-dependent and -independent pathways in E2F-mediated S phase induction. J Biol Chem. 2000;275:6337–6345. - PubMed
-
- Bahler J, Wu JQ, Longtine MS, Shah NG, McKenzie A, 3rd, Steever AB, Wach A, Philippsen P, Pringle JR. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast. 1998;14:943–951. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
