

Comparative Study
doi: 10.1101/gad.959102.
The zinc-finger proto-oncogene Gfi-1b is essential for development of the erythroid and megakaryocytic lineages
Affiliations
- PMID: 11825872
- PMCID: PMC155332
- DOI: 10.1101/gad.959102
Item in Clipboard
Comparative Study
The zinc-finger proto-oncogene Gfi-1b is essential for development of the erythroid and megakaryocytic lineages
Genes Dev.
.
Abstract
Gfi-1 and Gfi-1b are novel proto-oncogenes identified by retroviral insertional mutagenesis. By gene targeting, we establish that Gfi-1b is required for the development of two related blood lineages, erythroid and megakaryocytic, in mice. Gfi-1b(-/-) embryonic stem cells fail to contribute to red cells of adult chimeras. Gfi-1b(-/-) embryos exhibit delayed maturation of primitive erythrocytes and subsequently die with failure to produce definitive enucleated erythrocytes. The fetal liver of mutant mice contains erythroid and megakaryocytic precursors arrested in their development. Myelopoiesis is normal. Therefore, Gfi-1b is an essential transcriptional regulator of erythroid and megakaryocyte development.
Figures
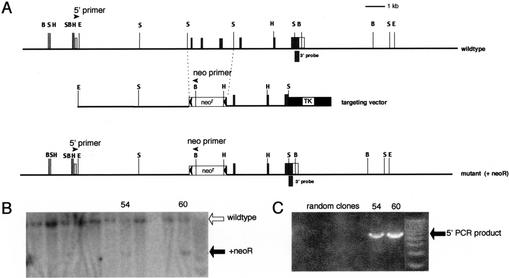
Targeted disruption of the mouse Gfi-1b gene. (A) Partial restriction map of the mouse Gfi-1b locus (top), the targeting vector (middle), and the expected targeted loci with the floxed neoR cassette (bottom). The 130-bp probe extending from the SacI site to the end of the Gfi-1b coding sequence on exon 7 used to detect appropriate 3′ integration of the targeting vector on Southern blots is indicated (3′ probe). The positions of the primers used to determine the 5′ integration by PCR are also indicated by arrowheads (5′ primer and neo primer, respectively). The Gfi-1b coding exons are indicated as shaded boxes, and the noncoding ones by open boxes. The floxed neoR cassette is indicated by an open box (neoR) flanked by arrowheads (loxP sites), and the TK cassette is shown as a solid black box. The restriction enzyme sites indicated in the map are BamHI (B), EcoRI (E), HindIII (H), and SacI (S). The sizes of the BamHI fragment detected by the 3′ probe in the wild-type and the mutant allele with the inserted neoR cassette are 12 kb and 5 kb, respectively. (B) Southern blot analysis of G148- and gancyclovir-resistant ES cell clones with the 3′ probe. Positions of the wild-type and mutant alleles (with neoR) are indicated by open and solid arrows, respectively. (C) PCR amplification of selected clones shown in B with the 5′ and neo primers, respectively. The PCR product indicative of the homologous recombination is indicated (5′ PCR product).
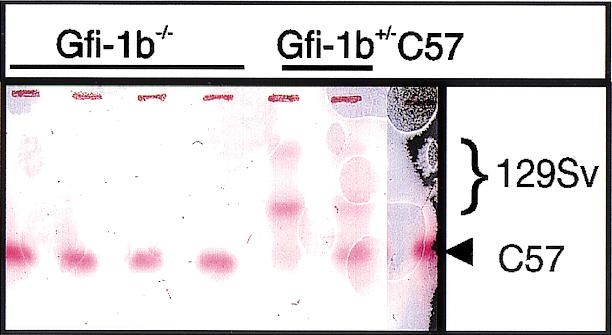
Gfi-1b−/− ES cells fail to contribute to adult red cell hemoglobin in chimeric mice. Hemoglobin electrophoresis of peripheral blood from four Gfi-1b−/− chimeric mice (lanes 1–4), two Gfi-1b+/− chimeric mice (lanes 5,6), and a wild-type C57Bl/6 mouse.
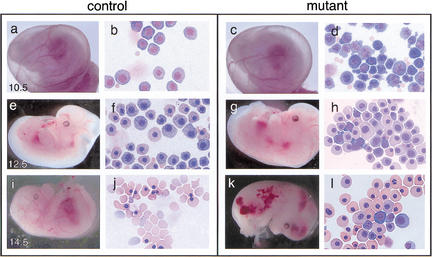
Control and Gfi-1b mutant embryos and peripheral blood at different gestational ages. Control (a,e,i) and mutant (c,g,k) embryos at E10.5, E12.5, and E14.5 and May–Grunwald–Giemsa stains of their corresponding yolk sac blood (b, f, and j, and d, h, and l, respectively). Gfi-1b−/− embryos show aberrant primitive erythropoiesis characterized by abnormal cell morphology (d) and delayed cellular maturation (h,l). Embryos die by E15 (k) from a failure of fetal liver erythropoiesis, resulting in the complete absence of definitive enucleated erythrocytes (l).

Gfi-1b−/− fetal livers show arrested definitive erythropoiesis. (A) Flow cytometry of E12.5 fetal livers. Forward (FSC-H) and side scatter (SSC-H) profiles of control (a) and mutant fetal livers (c). FACS profiles of gated (b,d) fetal liver cells stained with antibodies to c-kit and ter119. The majority of cells (60%–70%) from control livers are ter119hi and c-kit− (b), showing normal erythroid maturation. Cells from Gfi-1b−/− livers are either ter119− or ter119lo and c-kit+ (d). (B) Fetal liver cells from control embryos produce CFU-Es (day 3, a) and BFU-Es (day 7, b) when cultured in vitro with epo and KL. Gfi-1b−/− cells proliferate in epo and KL (e) but cannot mature into BFU-Es (e) or CFU-Es (d). Cells from BFU-E colonies of control fetal livers stain positively for benzidine (brown/black cells, c), but those from Gfi-1b−/− liver colonies do not (f).
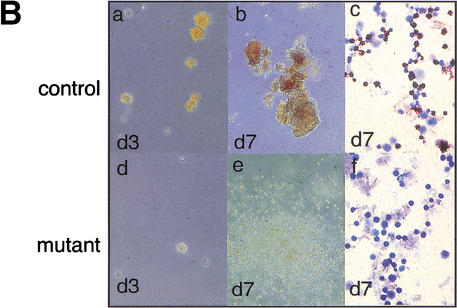
Gfi-1b−/− fetal livers show arrested definitive erythropoiesis. (A) Flow cytometry of E12.5 fetal livers. Forward (FSC-H) and side scatter (SSC-H) profiles of control (a) and mutant fetal livers (c). FACS profiles of gated (b,d) fetal liver cells stained with antibodies to c-kit and ter119. The majority of cells (60%–70%) from control livers are ter119hi and c-kit− (b), showing normal erythroid maturation. Cells from Gfi-1b−/− livers are either ter119− or ter119lo and c-kit+ (d). (B) Fetal liver cells from control embryos produce CFU-Es (day 3, a) and BFU-Es (day 7, b) when cultured in vitro with epo and KL. Gfi-1b−/− cells proliferate in epo and KL (e) but cannot mature into BFU-Es (e) or CFU-Es (d). Cells from BFU-E colonies of control fetal livers stain positively for benzidine (brown/black cells, c), but those from Gfi-1b−/− liver colonies do not (f).
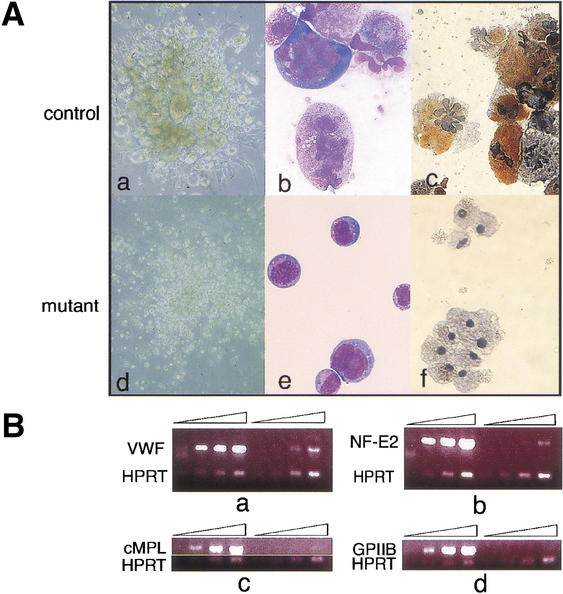
Gfi-1b−/− fetal livers show arrested megakaryopoiesis. (A) Colonies (a,d) and cells (b,c,e,f) from control (a–c) and Gfi-1b−/− (d–f) fetal liver cells grown in thrombopoietin (tpo). When cultured in tpo, fetal liver cells from wild-type livers give colonies with large megakaryocytes (a), whereas Gfi-1b−/− cells proliferate in tpo but do not differentiate into large megakaryocytes (d). The Gfi-1b−/− cells also do not show any nuclear (multilobulation) or cytoplasmic (granulation) maturation upon May–Grunwald–Giemsa staining (b vs. e) and are negative for acetylcholine esterase staining (c vs. f). (B) Semiquantitative RT–PCR of control (wild-type) and Gfi-1b−/− fetal liver cells cultured in tpo. Gfi-1b−/− cells have far fewer transcripts encoding markers of mature megakaryocytes relative to controls (a–d, lanes 5–8 vs. 1–4), for example, von-Willebrand factor (vWF in a), the transcription factor NF-E2 (b), the c-MPL receptor (c), and the surface glycoprotein IIb (d).
Similar articles
-
Erythroid expansion mediated by the Gfi-1B zinc finger protein: role in normal hematopoiesis.Blood. 2002 Oct 15;100(8):2769-77. doi: 10.1182/blood-2002-01-0182. Blood. 2002. PMID: 12351384
-
Distinct, strict requirements for Gfi-1b in adult bone marrow red cell and platelet generation.J Exp Med. 2014 May 5;211(5):909-27. doi: 10.1084/jem.20131065. Epub 2014 Apr 7. J Exp Med. 2014. PMID: 24711581 Free PMC article.
-
Gfi-1B controls human erythroid and megakaryocytic differentiation by regulating TGF-beta signaling at the bipotent erythro-megakaryocytic progenitor stage.Blood. 2010 Apr 8;115(14):2784-95. doi: 10.1182/blood-2009-09-241752. Epub 2010 Feb 2. Blood. 2010. PMID: 20124515
-
Gfi-1 oncoproteins in hematopoiesis.Hematology. 2003 Oct;8(5):339-44. doi: 10.1080/10245330310001612116. Hematology. 2003. PMID: 14530176 Review.
-
Ontogeny of erythropoiesis in the mammalian embryo.Curr Top Dev Biol. 2008;82:1-22. doi: 10.1016/S0070-2153(07)00001-4. Curr Top Dev Biol. 2008. PMID: 18282515 Review.
Cited by
-
Inherited platelet dysfunction and hematopoietic transcription factor mutations.Platelets. 2017 Jan;28(1):20-26. doi: 10.1080/09537104.2016.1203400. Epub 2016 Jul 27. Platelets. 2017. PMID: 27463948 Free PMC article. Review.
-
GATA-1 forms distinct activating and repressive complexes in erythroid cells.EMBO J. 2005 Jul 6;24(13):2354-66. doi: 10.1038/sj.emboj.7600702. Epub 2005 May 26. EMBO J. 2005. PMID: 15920471 Free PMC article.
-
Massively parallel sequencing identifies the gene Megf8 with ENU-induced mutation causing heterotaxy.Proc Natl Acad Sci U S A. 2009 Mar 3;106(9):3219-24. doi: 10.1073/pnas.0813400106. Epub 2009 Feb 13. Proc Natl Acad Sci U S A. 2009. PMID: 19218456 Free PMC article.
-
Selective dissociation between LSD1 and GFI1B by a LSD1 inhibitor NCD38 induces the activation of ERG super-enhancer in erythroleukemia cells.Oncotarget. 2018 Apr 20;9(30):21007-21021. doi: 10.18632/oncotarget.24774. eCollection 2018 Apr 20. Oncotarget. 2018. PMID: 29765516 Free PMC article.
-
Transcriptional repressors, corepressors and chromatin modifying enzymes in T cell development.Cytokine. 2011 Mar;53(3):271-81. doi: 10.1016/j.cyto.2010.11.013. Epub 2010 Dec 16. Cytokine. 2011. PMID: 21163671 Free PMC article. Review.
References
-
- Fuchs B, Wagner T, Rossel N, Antoine M, Beug H, Niessing J. Structure and erythroid cell-restricted expression of a chicken cDNA encoding a novel zinc finger protein of the Cys + His class. Gene. 1997;195:277–284. - PubMed
-
- Gao Y, Sun Y, Frank KM, Dikkes P, Fujiwara Y, Seidl KJ, Sekiguchi JM, Rathbun GA, Swat W, Wang J, et al. A critical role for DNA end-joining proteins in both lymphogenesis and neurogenesis. Cell. 1998;95:891–902. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases