

Chromatin assembly factor I and Hir proteins contribute to building functional kinetochores in S. cerevisiae
- PMID: 11782447
- PMCID: PMC155315
- DOI: 10.1101/gad.925302
Chromatin assembly factor I and Hir proteins contribute to building functional kinetochores in S. cerevisiae
Abstract
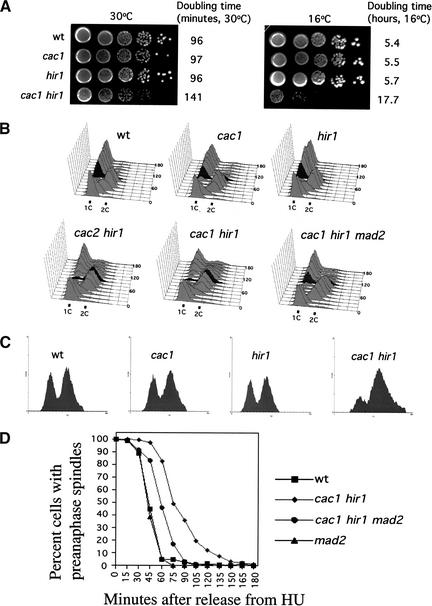
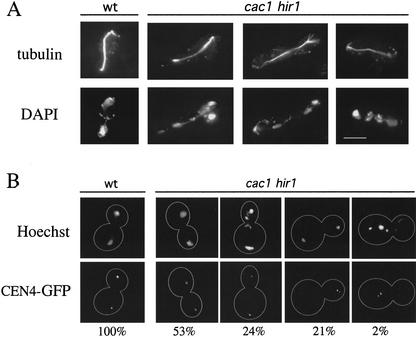
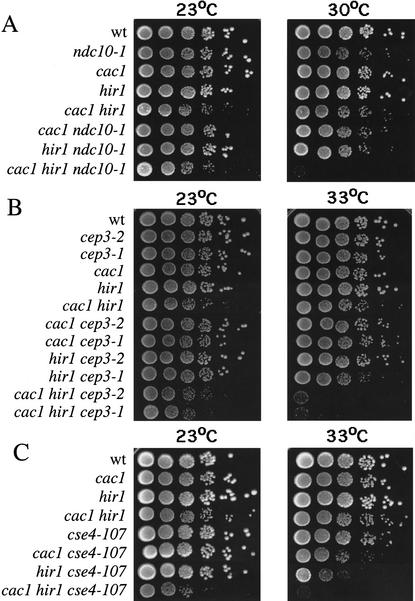
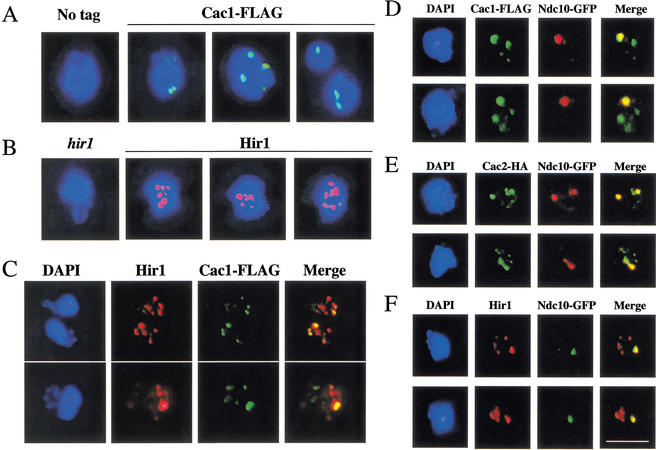
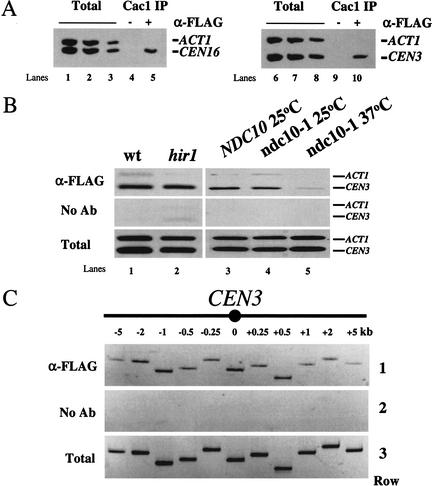
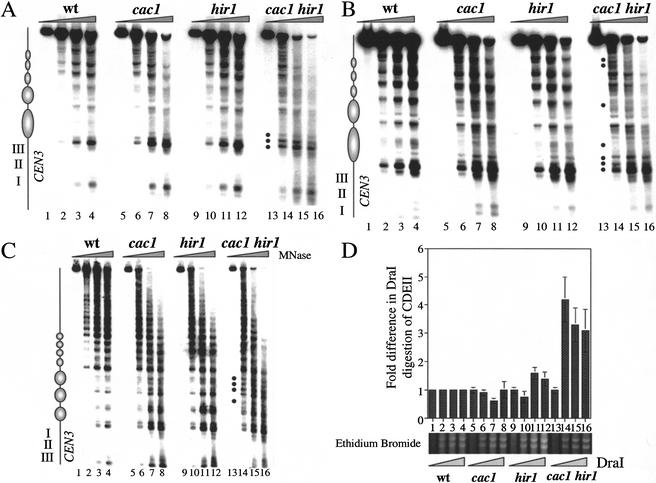
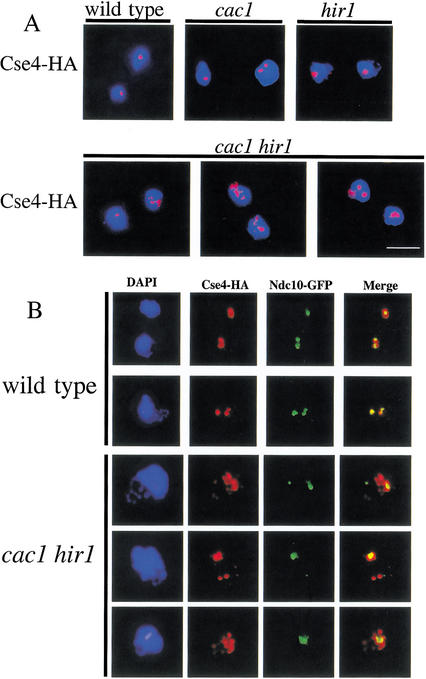
Budding yeast centromeres are comprised of approximately 125-bp DNA sequences that direct formation of the kinetochore, a specialized chromatin structure that mediates spindle attachment to chromosomes. We report here a novel role for the histone deposition complex chromatin assembly factor I (CAF-I) in building centromeric chromatin. The contribution of CAF-I to kinetochore function overlaps that of the Hir proteins, which have also been implicated in nucleosome formation and heterochromatic gene silencing. cacDelta hirDelta double mutant cells lacking both CAF-I and Hir proteins are delayed in anaphase entry in a spindle assembly checkpoint-dependent manner. Further, cacDelta and hirDelta deletions together cause increased rates of chromosome missegregation, genetic synergies with mutations in kinetochore protein genes, and alterations in centromeric chromatin structure. Finally, CAF-I subunits and Hir1 are enriched at centromeres, indicating that these proteins make a direct contribution to centromeric chromatin structures.
Figures
Similar articles
-
Chromatin assembly: the kinetochore connection.Curr Biol. 2002 Apr 2;12(7):R256-8. doi: 10.1016/s0960-9822(02)00786-8. Curr Biol. 2002. PMID: 11937044 Review.
-
Hir proteins are required for position-dependent gene silencing in Saccharomyces cerevisiae in the absence of chromatin assembly factor I.Mol Cell Biol. 1998 Aug;18(8):4793-806. doi: 10.1128/MCB.18.8.4793. Mol Cell Biol. 1998. PMID: 9671489 Free PMC article.
-
The budding yeast silencing protein Sir1 is a functional component of centromeric chromatin.Genes Dev. 2003 Oct 1;17(19):2356-61. doi: 10.1101/gad.1131103. Epub 2003 Sep 15. Genes Dev. 2003. PMID: 12975325 Free PMC article.
-
Yeast histone deposition protein Asf1p requires Hir proteins and PCNA for heterochromatic silencing.Curr Biol. 2001 Apr 3;11(7):463-73. doi: 10.1016/s0960-9822(01)00140-3. Curr Biol. 2001. PMID: 11412995
-
Structure, function, and regulation of budding yeast kinetochores.Annu Rev Cell Dev Biol. 2003;19:519-39. doi: 10.1146/annurev.cellbio.19.111301.155607. Annu Rev Cell Dev Biol. 2003. PMID: 14570580 Review.
Cited by
-
Histone H4 Facilitates the Proteolysis of the Budding Yeast CENP-ACse4 Centromeric Histone Variant.Genetics. 2017 Jan;205(1):113-124. doi: 10.1534/genetics.116.194027. Epub 2016 Oct 28. Genetics. 2017. PMID: 27794026 Free PMC article.
-
Suppressor analysis of a histone defect identifies a new function for the hda1 complex in chromosome segregation.Genetics. 2006 May;173(1):435-50. doi: 10.1534/genetics.105.050559. Epub 2006 Jan 16. Genetics. 2006. PMID: 16415367 Free PMC article.
-
Dysfunctional CAF-I reveals its role in cell cycle progression and differential regulation of gene silencing.Cell Cycle. 2019 Nov;18(22):3223-3236. doi: 10.1080/15384101.2019.1673100. Epub 2019 Sep 29. Cell Cycle. 2019. PMID: 31564230 Free PMC article.
-
A protein interaction map of the mitotic spindle.Mol Biol Cell. 2007 Oct;18(10):3800-9. doi: 10.1091/mbc.e07-06-0536. Epub 2007 Jul 18. Mol Biol Cell. 2007. PMID: 17634282 Free PMC article.
-
Silencing of chromatin assembly factor 1 in human cells leads to cell death and loss of chromatin assembly during DNA synthesis.Mol Cell Biol. 2004 Apr;24(7):2853-62. doi: 10.1128/MCB.24.7.2853-2862.2004. Mol Cell Biol. 2004. PMID: 15024074 Free PMC article.
References
-
- Allshire RC, Nimmo ER, Ekwall K, Javerzat JP, Cranston G. Mutations derepressing silent centromeric domains in fission yeast disrupt chromosome segregation. Genes & Dev. 1995;9:218–233. - PubMed
-
- Amon A. The spindle checkpoint. Curr Opin Genet Dev. 1999;9:69–75. - PubMed
-
- Blat Y, Kleckner N. Cohesins bind to preferential sites along yeast chromosome III, with differential regulation along arms versus the centric region. Cell. 1999;98:249–259. - PubMed
-
- Bloom KS, Carbon J. Yeast centromere DNA is in a unique and highly ordered structure in chromosomes and small circular minichromosomes. Cell. 1982;29:305–317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases