

Translation initiation control by heme-regulated eukaryotic initiation factor 2alpha kinase in erythroid cells under cytoplasmic stresses
- PMID: 11689689
- PMCID: PMC99965
- DOI: 10.1128/MCB.21.23.7971-7980.2001
Translation initiation control by heme-regulated eukaryotic initiation factor 2alpha kinase in erythroid cells under cytoplasmic stresses
Abstract
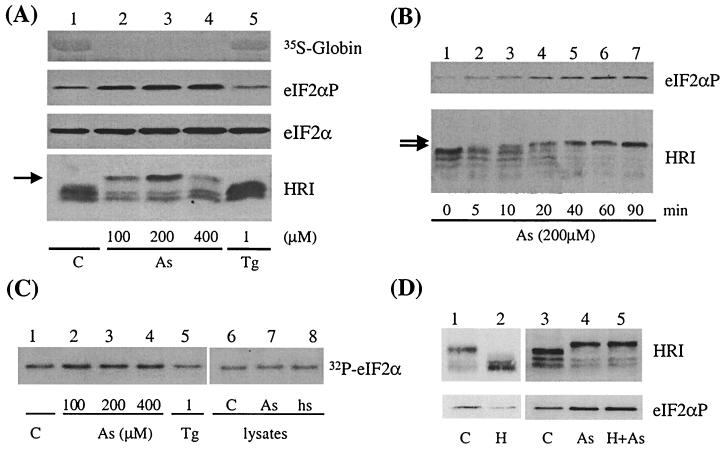
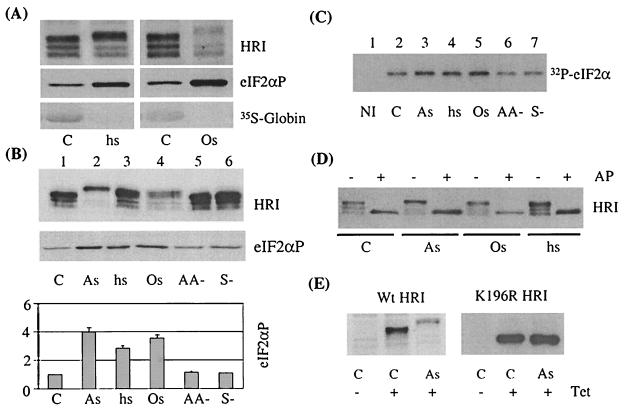
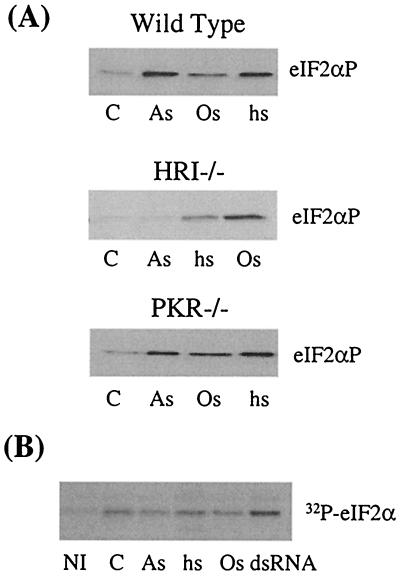
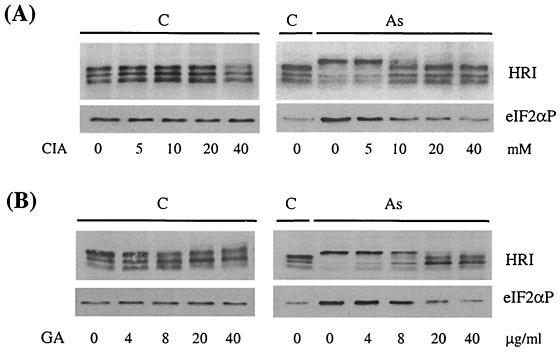
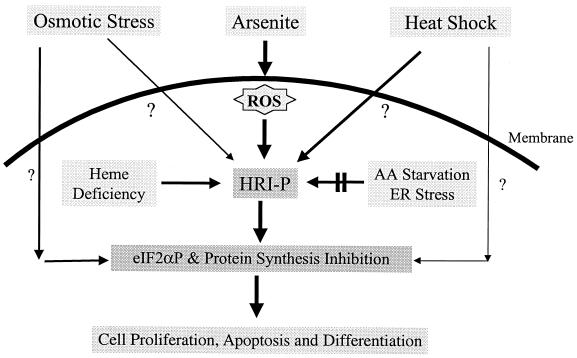
Cytoplasmic stresses, including heat shock, osmotic stress, and oxidative stress, cause rapid inhibition of protein synthesis in cells through phosphorylation of eukaryotic initiation factor 2alpha (eIF2alpha) by eIF2alpha kinases. We have investigated the role of heme-regulated inhibitor (HRI), a heme-regulated eIF2alpha kinase, in stress responses of erythroid cells. We have demonstrated that HRI in reticulocytes and fetal liver nucleated erythroid progenitors is activated by oxidative stress induced by arsenite, heat shock, and osmotic stress but not by endoplasmic reticulum stress or nutrient starvation. While autophosphorylation is essential for the activation of HRI, the phosphorylation status of HRI activated by different stresses is different. The contributions of HRI in various stress responses were assessed with the aid of HRI-null reticulocytes and fetal liver erythroid cells. HRI is the only eIF2alpha kinase activated by arsenite in erythroid cells, since HRI-null cells do not induce eIF2alpha phosphorylation upon arsenite treatment. HRI is also the major eIF2alpha kinase responsible for the increased eIF2alpha phosphorylation upon heat shock in erythroid cells. Activation of HRI by these stresses is independent of heme and requires the presence of intact cells. Both hsp90 and hsc70 are necessary for all stress-induced HRI activation. However, reactive oxygen species are involved only in HRI activation by arsenite. Our results provide evidence for a novel function of HRI in stress responses other than heme deficiency.
Figures


Similar articles
-
Evidence that Hsc70 negatively modulates the activation of the heme-regulated eIF-2alpha kinase in rabbit reticulocyte lysate.Eur J Biochem. 1998 Aug 1;255(3):552-62. doi: 10.1046/j.1432-1327.1998.2550552.x. Eur J Biochem. 1998. PMID: 9738893
-
A protective role of heme-regulated eIF2α kinase in cadmium-induced toxicity in erythroid cells.Food Chem Toxicol. 2013 Dec;62:880-91. doi: 10.1016/j.fct.2013.10.017. Epub 2013 Oct 22. Food Chem Toxicol. 2013. PMID: 24161693
-
Hsp90 mediates activation of the heme regulated eIF-2 alpha kinase during oxidative stress.Indian J Biochem Biophys. 2010 Apr;47(2):67-74. Indian J Biochem Biophys. 2010. PMID: 20521618
-
[Role of HRI in apoptosis resistance].Med Sci (Paris). 2014 Oct;30(10):882-8. doi: 10.1051/medsci/20143010015. Epub 2014 Oct 14. Med Sci (Paris). 2014. PMID: 25311023 Review. French.
-
Heme-regulated inhibitor: an overlooked eIF2α kinase in cancer investigations.Med Oncol. 2022 May 15;39(5):73. doi: 10.1007/s12032-022-01668-1. Med Oncol. 2022. PMID: 35568791 Review.
Cited by
-
Human Betacoronavirus OC43 Interferes with the Integrated Stress Response Pathway in Infected Cells.Viruses. 2024 Jan 31;16(2):212. doi: 10.3390/v16020212. Viruses. 2024. PMID: 38399988 Free PMC article.
-
Cellular Stress: Modulator of Regulated Cell Death.Biology (Basel). 2023 Aug 25;12(9):1172. doi: 10.3390/biology12091172. Biology (Basel). 2023. PMID: 37759572 Free PMC article. Review.
-
Stress and Liver Fibrogenesis: Understanding the Role and Regulation of Stress Response Pathways in Hepatic Stellate Cells.Am J Pathol. 2023 Oct;193(10):1363-1376. doi: 10.1016/j.ajpath.2023.06.006. Epub 2023 Jul 7. Am J Pathol. 2023. PMID: 37422148 Free PMC article. Review.
-
The integrated stress response is activated in the salivary glands of Sjögren's syndrome patients.Front Med (Lausanne). 2023 Mar 23;10:1118703. doi: 10.3389/fmed.2023.1118703. eCollection 2023. Front Med (Lausanne). 2023. PMID: 37035319 Free PMC article.
-
PPM1D suppresses p53-dependent transactivation and cell death by inhibiting the Integrated Stress Response.Nat Commun. 2022 Dec 1;13(1):7400. doi: 10.1038/s41467-022-35089-5. Nat Commun. 2022. PMID: 36456590 Free PMC article.
References
-
- Adler V, Yin Z, Tew K D, Ronai Z. Role of redox potential and reactive oxygen species in stress signaling. Oncogene. 1999;18:6104–6111. - PubMed
-
- Bauer, B. N., M. Rafie-Kolpin, L. Lu, A. Han, and J.-J. Chen. Multiple autophosphorylation is essential for the formation of the active and stable homodimer of heme-regulated eIF-2α kinase. Biochemistry, in press. - PubMed
-
- Berlanga J J, Santoyo J, DeHaro C. Characterization of a mammalian homolog of the GCN2 eukaryotic initiation factor 2alpha kinase. Eur J Biochem. 1999;265:754–762. - PubMed
-
- Brostrom C O, Brostrom M A. Regulation of translational initiation during cellular responses to stress. Prog Nucleic Acid Res Mol Biol. 1998;58:79–125. - PubMed
-
- Caplan A J. Hsp90's secrets unfold: new insights from structural and functional studies. Trends Cell Biol. 1999;9:262–268. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous