

RotundRacGAP functions with Ras during spermatogenesis and retinal differentiation in Drosophila melanogaster
- PMID: 11509670
- PMCID: PMC87355
- DOI: 10.1128/MCB.21.18.6280-6291.2001
RotundRacGAP functions with Ras during spermatogenesis and retinal differentiation in Drosophila melanogaster
Abstract
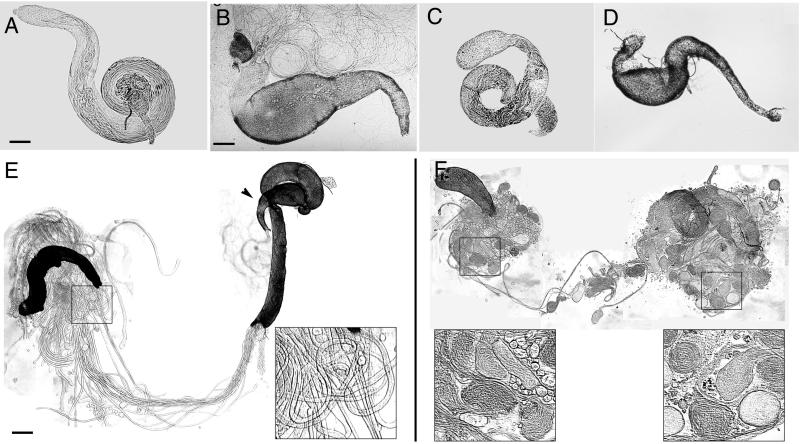
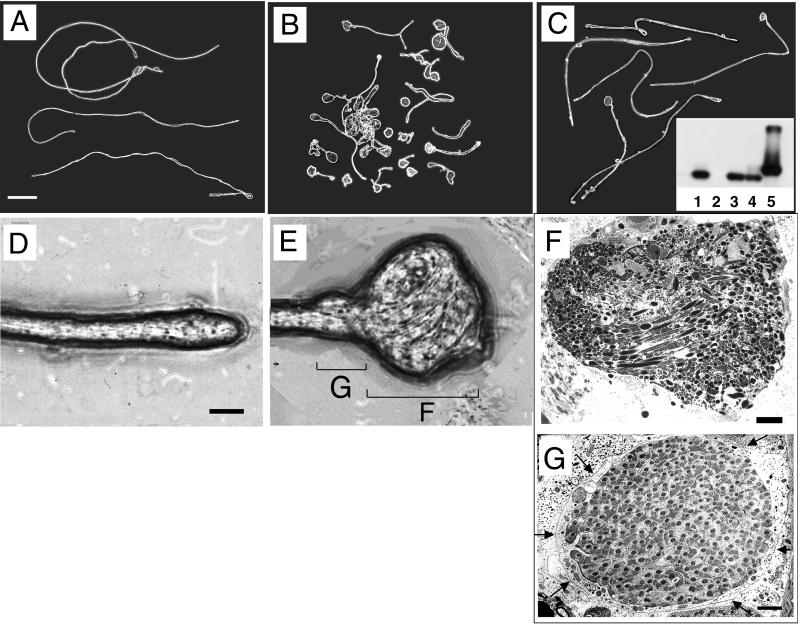
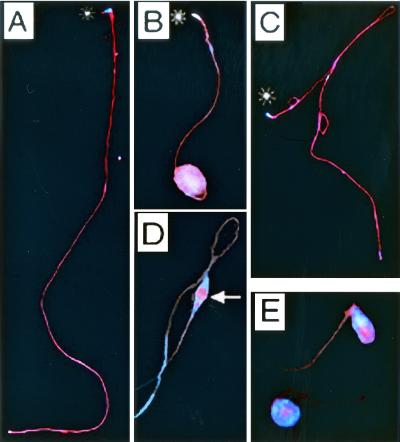
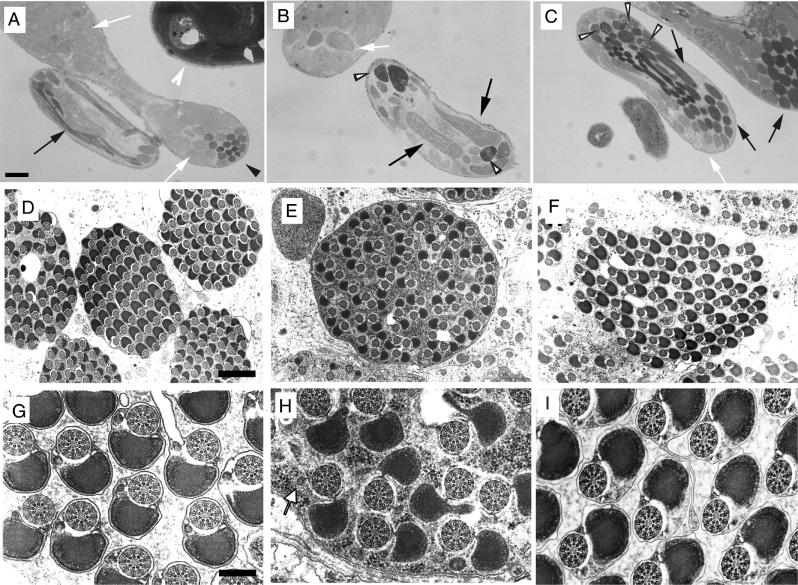
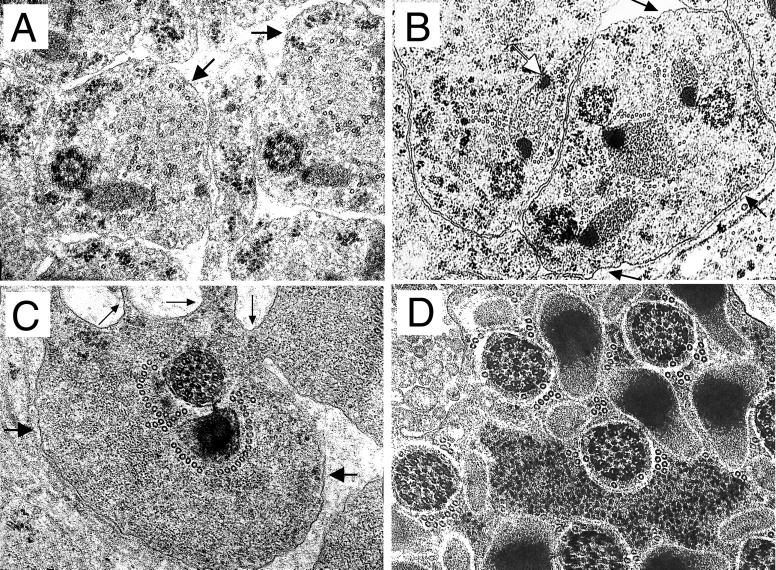
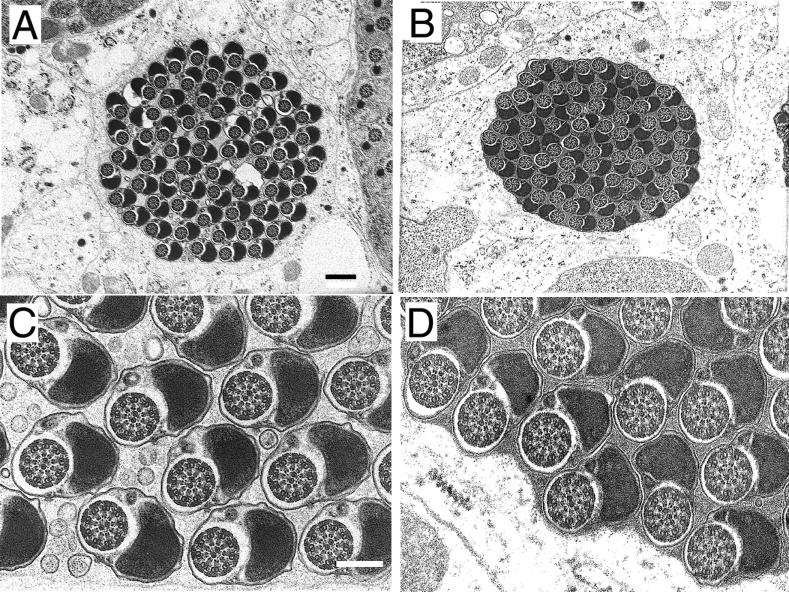
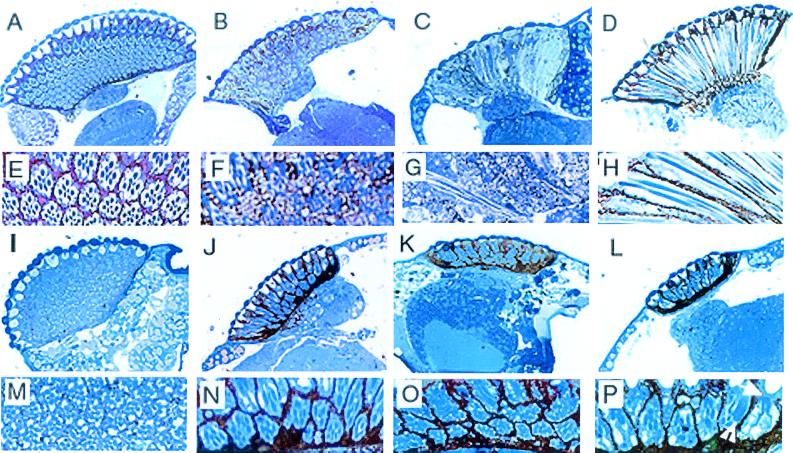
Our analysis of rotund (rn) null mutations in Drosophila melanogaster revealed that deletion of the rn locus affects both spermatid and retinal differentiation. In the male reproductive system, the absence of RnRacGAP induced small testes, empty seminal vesicles, short testicular cysts, reduced amounts of interspermatid membrane, the absence of individualization complexes, and incomplete mitochondrial condensation. Flagellar growth continued within the short rn null cysts to produce large bulbous terminations of intertwined mature flagella. Organization of the retina was also severely perturbed as evidenced by grossly misshapen ommatidia containing reduced numbers of photoreceptor and pigment cells. These morphological phenotypes were rescued by genomic rnRacGAP transgenes, demonstrating that RnRacGAP function is critical to spermatid and retinal differentiation. The testicular phenotypes were suppressed by heterozygous hypomorphic mutations in the Dras1 and drk genes, indicating cross talk between RacGAP-regulated signaling and that of the Ras pathway. The observed genetic interactions are consistent with a model in which Rac signaling is activated by Ras and negatively regulated by RnRacGAP during spermatid differentiation. RnRacGAP and Ras cross talk also operated during retinal differentiation; however, while the heterozygous hypomorphic drk mutation continued to act as a suppressor of the rn null mutation, the heterozygous hypomorphic Dras1 mutation induced novel retinal phenotypes.
Figures
Similar articles
-
Alternative splicing of the Drosophila melanogaster rotundRacGAP gene.Gene. 1996 Feb 12;168(2):135-41. doi: 10.1016/0378-1119(95)00747-4. Gene. 1996. PMID: 8654933
-
A screen for modifiers of RacGAP(84C) gain-of-function in the Drosophila eye revealed the LIM kinase Cdi/TESK1 as a downstream effector of Rac1 during spermatogenesis.J Cell Sci. 2004 Jun 1;117(Pt 13):2777-89. doi: 10.1242/jcs.01123. J Cell Sci. 2004. PMID: 15169836
-
Overexpression of RnRacGAP in Drosophila melanogaster deregulates cytoskeletal organisation in cellularising embryos and induces discrete imaginal phenotypes.Mech Dev. 1997 Jan;61(1-2):49-62. doi: 10.1016/s0925-4773(96)00619-3. Mech Dev. 1997. PMID: 9076677
-
[Universal intracellular transducer RAS and its role in the development of Drosophila].Ontogenez. 2013 Sep-Oct;44(5):331-40. Ontogenez. 2013. PMID: 25434202 Review. Russian.
-
The evolutionary diversity of insect retinal mosaics: common design principles and emerging molecular logic.Trends Genet. 2015 Jun;31(6):316-28. doi: 10.1016/j.tig.2015.04.006. Epub 2015 May 26. Trends Genet. 2015. PMID: 26025917 Free PMC article. Review.
Cited by
-
RacGap50C negatively regulates wingless pathway activity during Drosophila embryonic development.Genetics. 2005 Apr;169(4):2075-86. doi: 10.1534/genetics.104.039735. Epub 2005 Feb 3. Genetics. 2005. PMID: 15695356 Free PMC article.
-
A novel testicular RhoGAP-domain protein induces apoptosis.Biol Reprod. 2004 Dec;71(6):1980-90. doi: 10.1095/biolreprod.104.032805. Epub 2004 Aug 11. Biol Reprod. 2004. PMID: 15306557 Free PMC article.
-
Separating from the pack: Molecular mechanisms of Drosophila spermatid individualization.Spermatogenesis. 2015 May 21;5(2):e1041345. doi: 10.1080/21565562.2015.1041345. eCollection 2015 May-Aug. Spermatogenesis. 2015. PMID: 26413413 Free PMC article. No abstract available.
-
The role of Drosophila Merlin in spermatogenesis.BMC Cell Biol. 2008 Jan 10;9:1. doi: 10.1186/1471-2121-9-1. BMC Cell Biol. 2008. PMID: 18186933 Free PMC article.
-
Rho family GTPase Rnd2 interacts and co-localizes with MgcRacGAP in male germ cells.Biochem J. 2003 May 15;372(Pt 1):105-12. doi: 10.1042/BJ20021652. Biochem J. 2003. PMID: 12590651 Free PMC article.
References
-
- Agnel M, Kerridge S, Vola C, Griffin-Shea R. Two transcripts from the rotund region of Drosophila show similar positional specificities in imaginal disc tissues. Genes Dev. 1989;3:85–95. - PubMed
-
- Agnel M, Roder L, Griffin-Shea R, Vola C. The spatial expression of Drosophila rotund gene reveals that the imaginal discs are organized in domains along the proximal-distal axis. Roux's Arch Dev Biol. 1992;201:284–295. - PubMed
-
- Ashburner M. Drosophila: a laboratory handbook. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1989. p. 198.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous