

Induction of caspase-dependent apoptosis in cultured cells by the avian coronavirus infectious bronchitis virus
- PMID: 11413307
- PMCID: PMC114363
- DOI: 10.1128/JVI.75.14.6402-6409.2001
Induction of caspase-dependent apoptosis in cultured cells by the avian coronavirus infectious bronchitis virus
Abstract
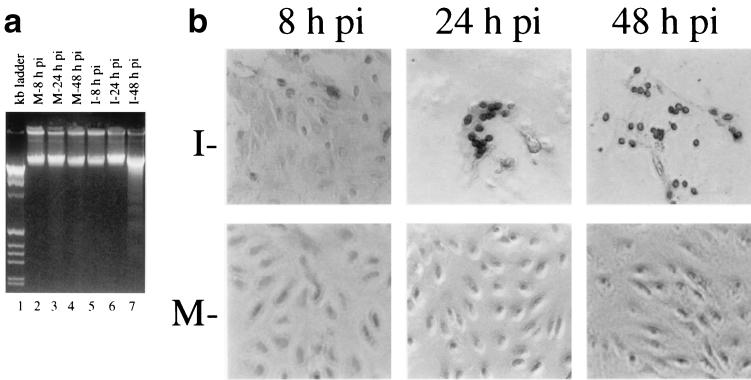
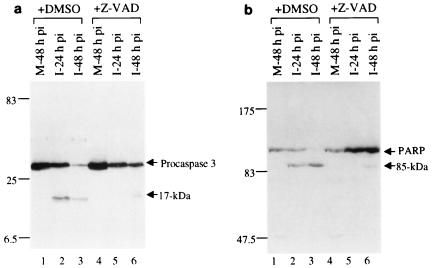
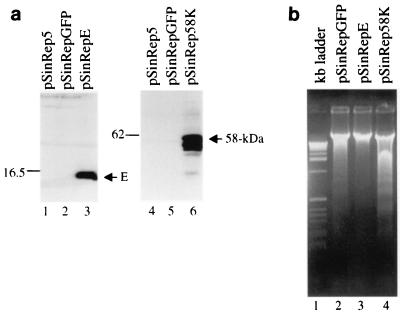
Avian coronavirus infectious bronchitis virus (IBV) is the causative agent of chicken infectious bronchitis, an acute, highly contagious viral respiratory disease. Replication of IBV in Vero cells causes extensive cytopathic effects (CPE), leading to destruction of the entire monolayer and the death of infected cells. In this study, we investigated the cell death processes during acute IBV infection and the underlying mechanisms. The results show that both necrosis and apoptosis may contribute to the death of infected cells in lytic IBV infection. Caspase-dependent apoptosis, as characterized by chromosomal condensation, DNA fragmentation, caspase-3 activation, and poly(ADP-ribose) polymerase degradation, was detected in IBV-infected Vero cells. Addition of the general caspase inhibitor z-VAD-FMK to the culture media showed inhibition of the hallmarks of apoptosis and increase of the release of virus to the culture media at 16 h postinfection. However, neither the necrotic process nor the productive replication of IBV in Vero cells was severely affected by the inhibition of apoptosis. Screening of 11 IBV-encoded proteins suggested that a 58-kDa mature cleavage product could induce apoptotic changes in cells transiently expressing the protein. This study adds one more example to the growing list of animal viruses that induce apoptosis during their replication cycles.
Figures


Similar articles
-
Infectious Bronchitis Virus Infection Induces Apoptosis during Replication in Chicken Macrophage HD11 Cells.Viruses. 2017 Jul 26;9(8):198. doi: 10.3390/v9080198. Viruses. 2017. PMID: 28933760 Free PMC article.
-
In vitro and in vivo effects of Houttuynia cordata on infectious bronchitis virus.Avian Pathol. 2011 Oct;40(5):491-8. doi: 10.1080/03079457.2011.605107. Epub 2011 Aug 18. Avian Pathol. 2011. PMID: 21848486
-
Up-regulation of Mcl-1 and Bak by coronavirus infection of human, avian and animal cells modulates apoptosis and viral replication.PLoS One. 2012;7(1):e30191. doi: 10.1371/journal.pone.0030191. Epub 2012 Jan 11. PLoS One. 2012. PMID: 22253918 Free PMC article.
-
Effects of a broad-spectrum caspase inhibitor, Z-VAD(OMe)-FMK, on viral hemorrhagic septicemia virus (VHSV) infection-mediated apoptosis and viral replication.Fish Shellfish Immunol. 2016 Apr;51:41-45. doi: 10.1016/j.fsi.2016.02.021. Epub 2016 Feb 17. Fish Shellfish Immunol. 2016. PMID: 26899629
-
Caspase cleavage of viral proteins, another way for viruses to make the best of apoptosis.Cell Death Dis. 2012 Mar 8;3(3):e277. doi: 10.1038/cddis.2012.18. Cell Death Dis. 2012. PMID: 22402601 Free PMC article. Review.
Cited by
-
Apoptosis Enhances the Replication of Human Coronavirus OC43.Viruses. 2021 Nov 1;13(11):2199. doi: 10.3390/v13112199. Viruses. 2021. PMID: 34835005 Free PMC article.
-
Equine torovirus (BEV) induces caspase-mediated apoptosis in infected cells.PLoS One. 2011;6(6):e20972. doi: 10.1371/journal.pone.0020972. Epub 2011 Jun 15. PLoS One. 2011. PMID: 21698249 Free PMC article.
-
Analysis of proteins that interact with nucleocapsid protein of SARS-CoV using 15-mer phage-displayed library.Chin Sci Bull. 2007;52(15):2072-2080. doi: 10.1007/s11434-007-0303-0. Chin Sci Bull. 2007. PMID: 32214725 Free PMC article.
-
Porcine reproductive and respiratory syndrome virus induces apoptosis through a mitochondria-mediated pathway.Virology. 2007 Sep 1;365(2):419-34. doi: 10.1016/j.virol.2007.04.001. Epub 2007 May 8. Virology. 2007. PMID: 17488647 Free PMC article.
-
Regulation of the ER Stress Response by the Ion Channel Activity of the Infectious Bronchitis Coronavirus Envelope Protein Modulates Virion Release, Apoptosis, Viral Fitness, and Pathogenesis.Front Microbiol. 2020 Jan 24;10:3022. doi: 10.3389/fmicb.2019.03022. eCollection 2019. Front Microbiol. 2020. PMID: 32038520 Free PMC article.
References
-
- Agol V I, Belov G A, Bienz K, Egger D, Kolesnikova M S, Raikhlin N T, Romanova L I, Smirnova E A, Tolskaya E A. Two types of death of poliovirus-infected cell: caspase involvement in the apoptosis but not cytopathic effect. Virology. 1998;252:343–353. - PubMed
-
- Arends M, Wellie A. Apoptosis: mechanism and roles in pathology. Int Rev Exp Pathol. 1991;32:223–254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
