

Mitogen-activated protein kinase phosphatase is required for genotoxic stress relief in Arabidopsis
- PMID: 11274055
- PMCID: PMC312655
- DOI: 10.1101/gad.192601
Mitogen-activated protein kinase phosphatase is required for genotoxic stress relief in Arabidopsis
Abstract
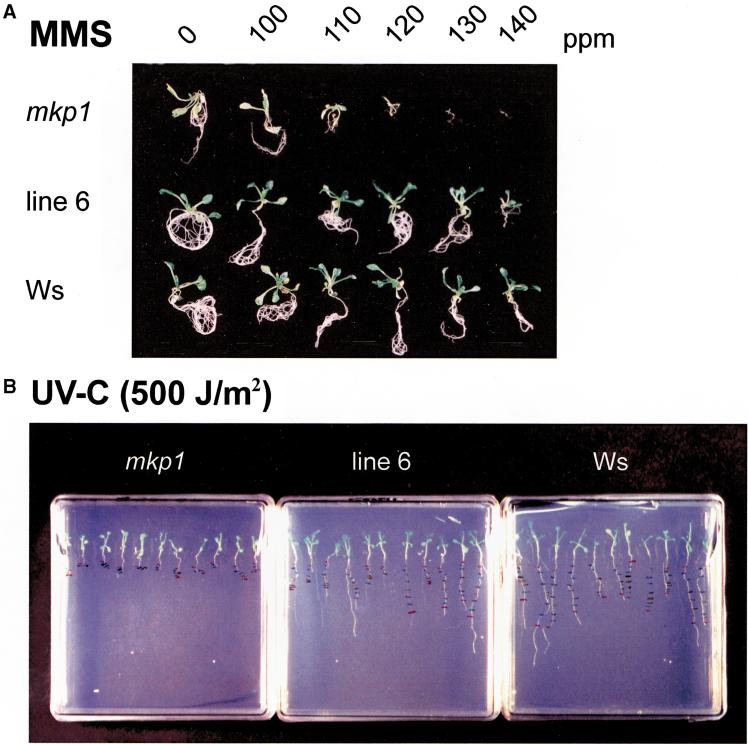
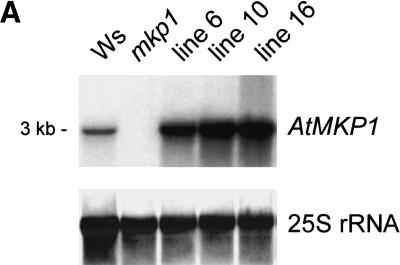
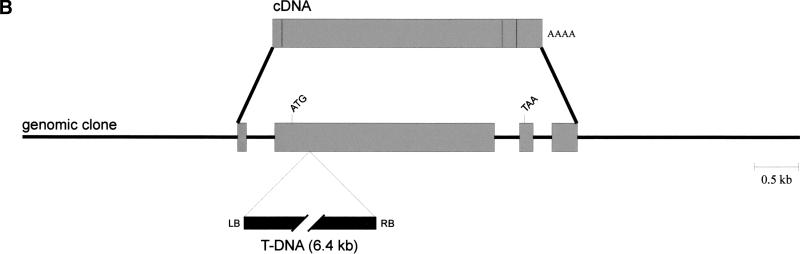
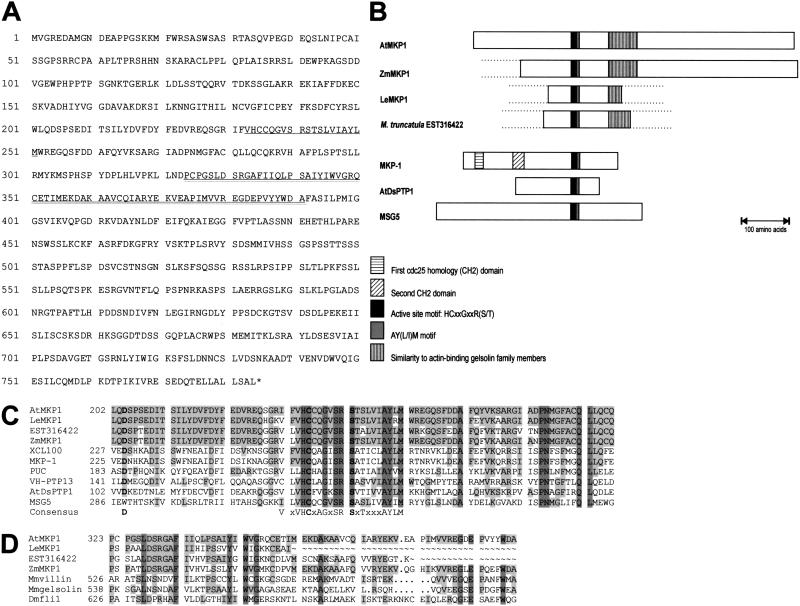
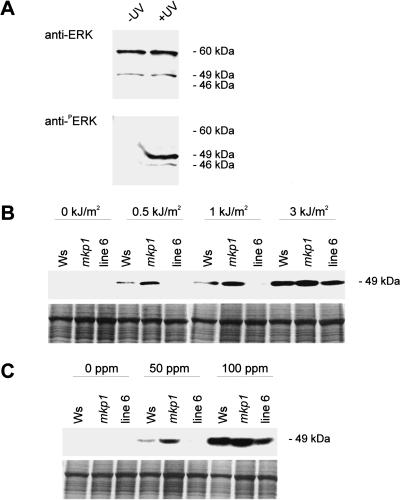
Genotoxic stress activates complex cellular responses allowing for the repair of DNA damage and proper cell recovery. Although plants are exposed constantly to increasing solar UV irradiation, the signaling cascades activated by genotoxic environments are largely unknown. We have identified an Arabidopsis mutant (mkp1) hypersensitive to genotoxic stress treatments (UV-C and methyl methanesulphonate) due to disruption of a gene that encodes an Arabidopsis homolog of mitogen-activated protein kinase phosphatase (AtMKP1). Growth of the mkp1 mutant under standard conditions is indistinguishable from wild type, indicating a stress-specific function of AtMKP1. MAP kinase phosphatases (MKPs), the potent inactivators of MAP kinases, are considered important regulators of MAP kinase signaling. Although biochemical data from mammalian cell cultures suggests an involvement of MKPs in cellular stress responses, there is no in vivo genetic support for this view in any multicellular organism. The genetic and biochemical data presented here imply a central role for a MAP kinase cascade in genotoxic stress signaling in plants and indicate AtMKP1 to be a crucial regulator of the MAP kinase activity in vivo, determining the outcome of the cellular reaction and the level of genotoxic resistance.
Figures
Similar articles
-
Phosphorylation and stabilization of Arabidopsis MAP kinase phosphatase 1 in response to UV-B stress.J Biol Chem. 2013 Jan 4;288(1):480-6. doi: 10.1074/jbc.M112.434654. Epub 2012 Nov 27. J Biol Chem. 2013. PMID: 23188831 Free PMC article.
-
Distinct regulation of salinity and genotoxic stress responses by Arabidopsis MAP kinase phosphatase 1.EMBO J. 2002 Dec 2;21(23):6483-93. doi: 10.1093/emboj/cdf646. EMBO J. 2002. PMID: 12456655 Free PMC article.
-
Arabidopsis MAP kinase phosphatase 1 and its target MAP kinases 3 and 6 antagonistically determine UV-B stress tolerance, independent of the UVR8 photoreceptor pathway.Plant J. 2011 Nov;68(4):727-37. doi: 10.1111/j.1365-313X.2011.04725.x. Epub 2011 Sep 13. Plant J. 2011. PMID: 21790814
-
Signaling through MAP kinase networks in plants.Arch Biochem Biophys. 2006 Aug 1;452(1):55-68. doi: 10.1016/j.abb.2006.05.001. Epub 2006 May 24. Arch Biochem Biophys. 2006. PMID: 16806044 Review.
-
Mitogen-activated protein kinase cascades in signaling plant growth and development.Trends Plant Sci. 2015 Jan;20(1):56-64. doi: 10.1016/j.tplants.2014.10.001. Epub 2014 Oct 22. Trends Plant Sci. 2015. PMID: 25457109 Review.
Cited by
-
TMKP1 is a novel wheat stress responsive MAP Kinase phosphatase localized in the nucleus.Plant Mol Biol. 2010 Jun;73(3):325-38. doi: 10.1007/s11103-010-9617-4. Epub 2010 Mar 4. Plant Mol Biol. 2010. PMID: 20204675
-
MKP1 acts as a key modulator of stomatal development.Plant Signal Behav. 2019;14(7):1604017. doi: 10.1080/15592324.2019.1604017. Epub 2019 Apr 13. Plant Signal Behav. 2019. PMID: 30983545 Free PMC article.
-
Cell signaling during cold, drought, and salt stress.Plant Cell. 2002;14 Suppl(Suppl):S165-83. doi: 10.1105/tpc.000596. Plant Cell. 2002. PMID: 12045276 Free PMC article. Review. No abstract available.
-
The PP2C-type phosphatase AP2C1, which negatively regulates MPK4 and MPK6, modulates innate immunity, jasmonic acid, and ethylene levels in Arabidopsis.Plant Cell. 2007 Jul;19(7):2213-24. doi: 10.1105/tpc.106.049585. Epub 2007 Jul 13. Plant Cell. 2007. PMID: 17630279 Free PMC article.
-
BRU1, a novel link between responses to DNA damage and epigenetic gene silencing in Arabidopsis.Genes Dev. 2004 Apr 1;18(7):782-93. doi: 10.1101/gad.295404. Genes Dev. 2004. PMID: 15082530 Free PMC article.
References
-
- Agarwal ML, Taylor WR, Chernov MV, Chernova OB, Stark GR. The p53 network. J Biol Chem. 1998;273:1–4. - PubMed
-
- Albinsky D, Masson JE, Bogucki A, Afsar K, Vass I, Nagy F, Paszkowski J. Plant responses to genotoxic stress are linked to an ABA/salinity signaling pathway. Plant J. 1999;17:73–82.
-
- Bechtold N, Ellis J, Pelletier G. In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C R Acad Sci Paris, Life Sci. 1993;316:1194–1199.
-
- Bouchez D, Vittoroso P, Courtial B, Camilleri C. Kanamycin rescue: A simple technique for the recovery of T-DNA flanking sequences. Plant Mol Biol Rep. 1996;14:115–123.
-
- Britt AB. DNA damage and repair in plants. Annu Rev Plant Physiol Plant Mol Biol. 1996;47:75–100. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous