

The tyrosine phosphatase SHP-2 is required for sustained activation of extracellular signal-regulated kinase and epithelial morphogenesis downstream from the met receptor tyrosine kinase
- PMID: 11046147
- PMCID: PMC102157
- DOI: 10.1128/MCB.20.22.8513-8525.2000
The tyrosine phosphatase SHP-2 is required for sustained activation of extracellular signal-regulated kinase and epithelial morphogenesis downstream from the met receptor tyrosine kinase
Abstract
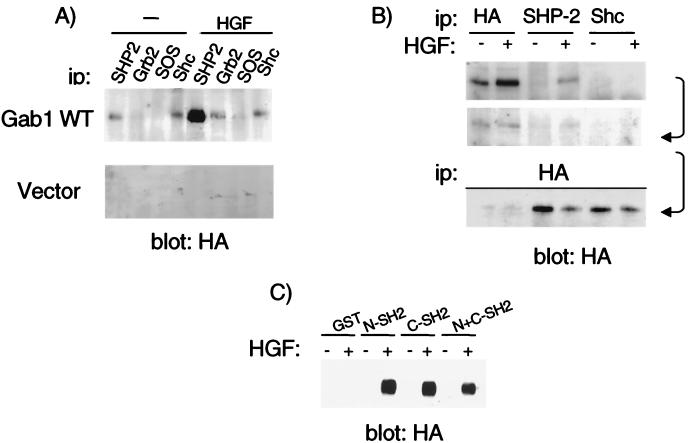
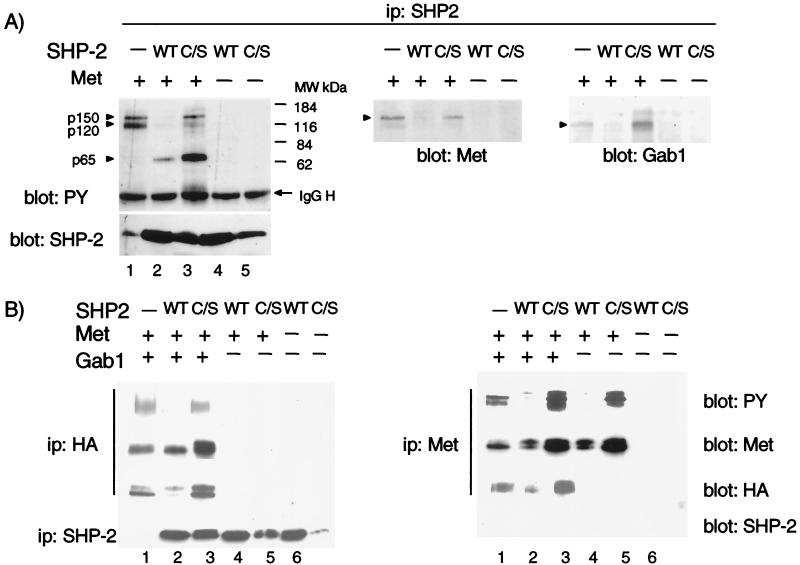
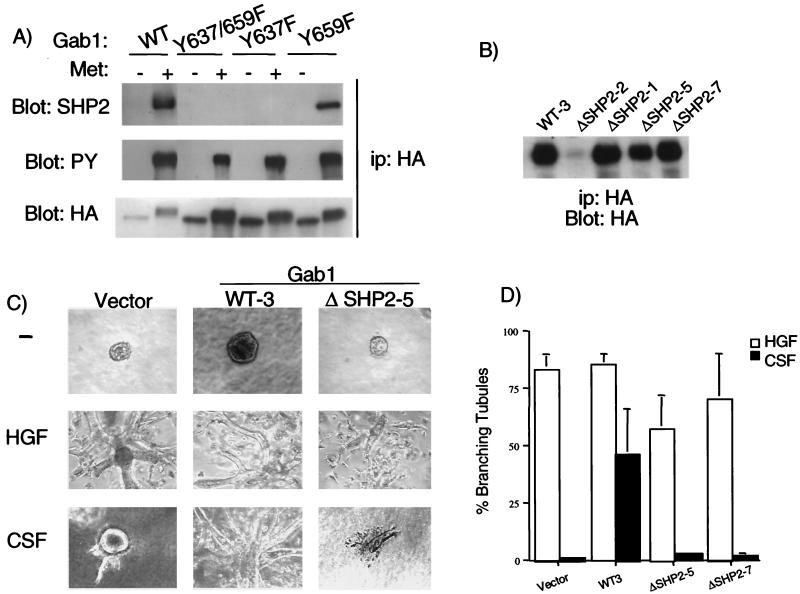
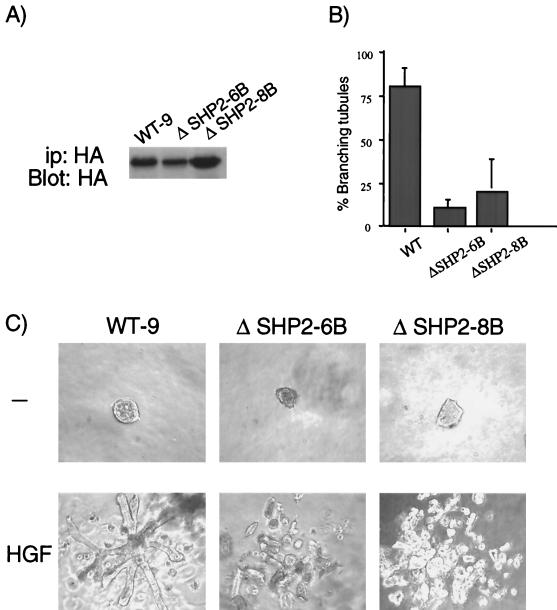
Epithelial morphogenesis is critical during development and wound healing, and alterations in this program contribute to neoplasia. Met, the hepatocyte growth factor (HGF) receptor, promotes a morphogenic program in epithelial cell lines in matrix cultures. Previous studies have identified Gab1, the major phosphorylated protein following Met activation, as important for the morphogenic response. Gab1 is a docking protein that couples the Met receptor with multiple signaling proteins, including phosphatidylinositol-3 kinase, phospholipase Cgamma, the adapter protein Crk, and the tyrosine specific phosphatase SHP-2. HGF induces sustained phosphorylation of Gab1 and sustained activation of extracellular signal-regulated kinase (Erk) in epithelial Madin-Darby canine kidney cells. In contrast, epidermal growth factor fails to promote a morphogenic program and induces transient Gab1 phosphorylation and Erk activation. To elucidate the Gab1-dependent signals required for epithelial morphogenesis, we undertook a structure-function approach and demonstrate that association of Gab1 with the tyrosine phosphatase SHP-2 is required for sustained Erk activation and for epithelial morphogenesis downstream from the Met receptor. Epithelial cells expressing a Gab1 mutant protein unable to recruit SHP-2 elicit a transient activation of Erk in response to HGF. Moreover, SHP-2 catalytic activity is required, since the expression of a catalytically inactive SHP-2 mutant, C/S, abrogates sustained activation of Erk and epithelial morphogenesis by the Met receptor. These data identify SHP-2 as a positive modulator of Erk activity and epithelial morphogenesis downstream from the Met receptor.
Figures

Similar articles
-
Role of the Grb2-associated binder 1/SHP-2 interaction in cell growth and transformation.Cancer Res. 2004 Mar 15;64(6):2007-15. doi: 10.1158/0008-5472.can-03-2886. Cancer Res. 2004. PMID: 15026337
-
The Gab1 PH domain is required for localization of Gab1 at sites of cell-cell contact and epithelial morphogenesis downstream from the met receptor tyrosine kinase.Mol Cell Biol. 1999 Mar;19(3):1784-99. doi: 10.1128/MCB.19.3.1784. Mol Cell Biol. 1999. PMID: 10022866 Free PMC article.
-
Distinct domains in the SHP-2 phosphatase differentially regulate epidermal growth factor receptor/NF-kappaB activation through Gab1 in glioblastoma cells.Mol Cell Biol. 2004 Jan;24(2):823-36. doi: 10.1128/MCB.24.2.823-836.2004. Mol Cell Biol. 2004. PMID: 14701753 Free PMC article.
-
Gab1 acts as an adapter molecule linking the cytokine receptor gp130 to ERK mitogen-activated protein kinase.Mol Cell Biol. 1998 Jul;18(7):4109-17. doi: 10.1128/MCB.18.7.4109. Mol Cell Biol. 1998. PMID: 9632795 Free PMC article.
-
The multisubstrate adapter Gab1 regulates hepatocyte growth factor (scatter factor)-c-Met signaling for cell survival and DNA repair.Mol Cell Biol. 2001 Aug;21(15):4968-84. doi: 10.1128/MCB.21.15.4968-4984.2001. Mol Cell Biol. 2001. PMID: 11438654 Free PMC article.
Cited by
-
Novel role for SHP-2 in nutrient-responsive control of S6 kinase 1 signaling.Mol Cell Biol. 2013 Jan;33(2):293-306. doi: 10.1128/MCB.01285-12. Epub 2012 Nov 5. Mol Cell Biol. 2013. PMID: 23129808 Free PMC article.
-
Salicylic acid based small molecule inhibitor for the oncogenic Src homology-2 domain containing protein tyrosine phosphatase-2 (SHP2).J Med Chem. 2010 Mar 25;53(6):2482-93. doi: 10.1021/jm901645u. J Med Chem. 2010. PMID: 20170098 Free PMC article.
-
The adaptor protein soc-1/Gab1 modifies growth factor receptor output in Caenorhabditis elegans.Genetics. 2006 May;173(1):163-75. doi: 10.1534/genetics.106.055822. Epub 2006 Mar 17. Genetics. 2006. PMID: 16547100 Free PMC article.
-
Dysregulation of the Met pathway in non-small cell lung cancer: implications for drug targeting and resistance.Transl Lung Cancer Res. 2015 Jun;4(3):242-52. doi: 10.3978/j.issn.2218-6751.2015.03.05. Transl Lung Cancer Res. 2015. PMID: 26207212 Free PMC article. Review.
-
Met receptor tyrosine kinase signals through a cortactin-Gab1 scaffold complex, to mediate invadopodia.J Cell Sci. 2012 Jun 15;125(Pt 12):2940-53. doi: 10.1242/jcs.100834. Epub 2012 Feb 24. J Cell Sci. 2012. PMID: 22366451 Free PMC article.
References
-
- Allard J D, Chang H C, Herbst R, McNeill H, Simon M A. The SH2-containing tyrosine phosphatase corkscrew is required during signaling by sevenless, Ras and Raf. Development. 1996;122:1137–1146. - PubMed
-
- Allard J D, Herbst R, Carroll P M, Simon M A. Mutational analysis of the SRC homology 2 domain protein-tyrosine phosphatase Corkscrew. J Biol Chem. 1998;273:13129–13135. - PubMed
-
- Barford D, Neel B G. Revealing mechanisms for SH2 domain-mediated regulation of the protein tyrosine phosphatase SHP-2. Structure. 1998;6:249–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous