

Role of the Rous sarcoma virus p10 domain in shape determination of gag virus-like particles assembled in vitro and within Escherichia coli
- PMID: 11024160
- PMCID: PMC102070
- DOI: 10.1128/jvi.74.21.10260-10268.2000
Role of the Rous sarcoma virus p10 domain in shape determination of gag virus-like particles assembled in vitro and within Escherichia coli
Abstract
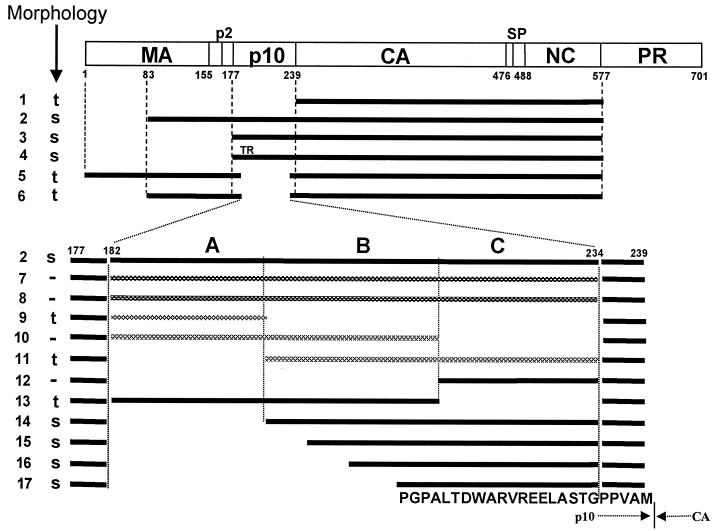
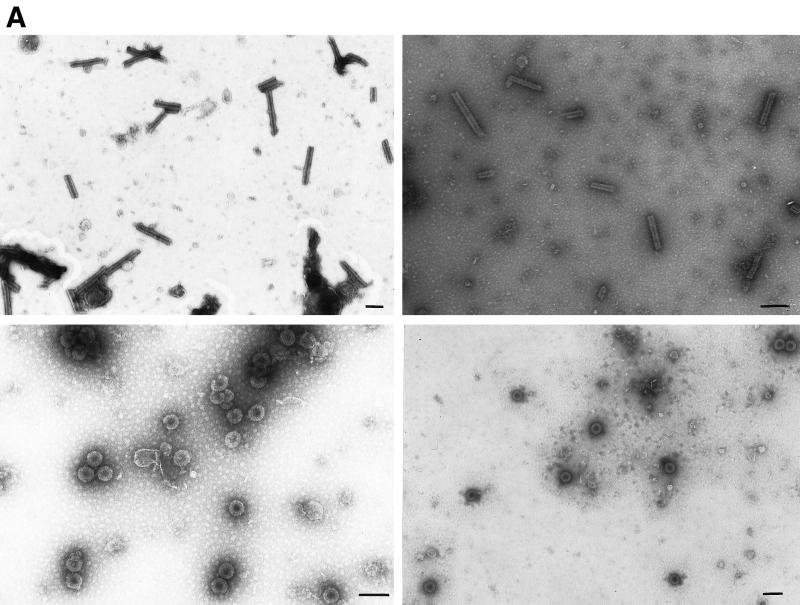
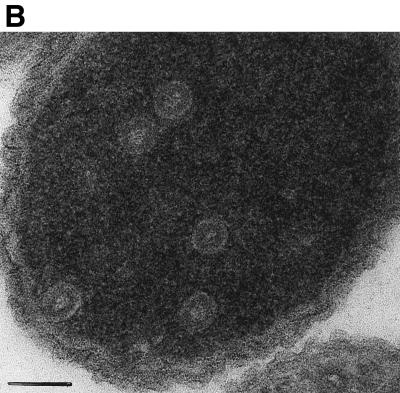
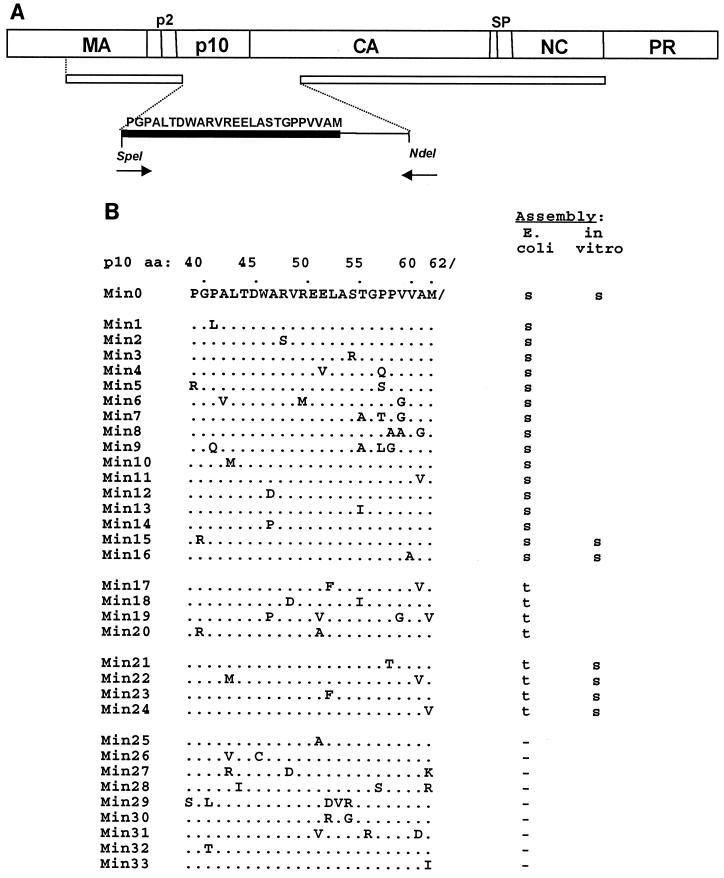
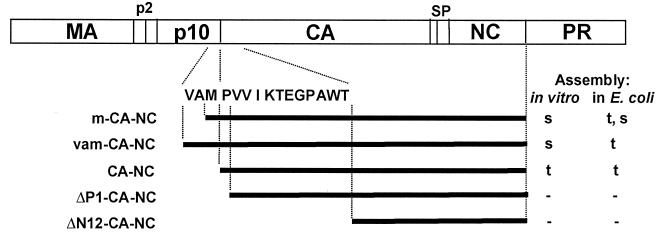
Purified retrovirus Gag proteins can assemble in vitro into virus-like particles (VLPs) in the presence of RNA. It was shown previously that a Rous sarcoma virus Gag protein missing only the protease domain forms spherical particles resembling immature virions lacking a membrane but that a similar protein missing the p10 domain forms tubular particles. Thus, p10 plays a role in spherical particle formation. To further study this shape-determining function, we dissected the p10 domain by mutagenesis and examined VLPs assembled within Escherichia coli or assembled in vitro from purified proteins. The results identified a minimal contiguous segment of 25 amino acid residues at the C terminus of p10 that is sufficient to restore efficient spherical assembly to a p10 deletion mutant. Random and site-directed mutations were introduced into this segment of polypeptide, and the shapes of particles formed in E. coli were examined in crude extracts by electron microscopy. Three phenotypes were observed: tubular morphology, spherical morphology, or no regular structure. While the particle morphology visualized in crude extracts generally was the same as that visualized for purified proteins, some tubular mutants scored as spherical when tested as purified proteins, suggesting that a cellular factor may also play a role in shape determination. We also examined the assembly properties of smaller Gag proteins consisting of the capsid protein-nucleocapsid protein (CA-NC) domains with short N-terminal extensions or deletions. Addition of one or three residues allowed CA-NC to form spheres instead of tubes in vitro, but the efficiency of assembly was extremely low. Deletion of the N-terminal residue(s) abrogated assembly. Taken together, these results imply that the N terminus of CA and the adjacent upstream 25 residues play an important role in the polymerization of the Gag protein.
Figures
Similar articles
-
Assembly and architecture of HIV.Adv Exp Med Biol. 2012;726:441-65. doi: 10.1007/978-1-4614-0980-9_20. Adv Exp Med Biol. 2012. PMID: 22297526 Free PMC article. Review.
-
Characterization of Rous sarcoma virus Gag particles assembled in vitro.J Virol. 2001 Mar;75(6):2753-64. doi: 10.1128/JVI.75.6.2753-2764.2001. J Virol. 2001. PMID: 11222698 Free PMC article.
-
In vitro assembly of virus-like particles with Rous sarcoma virus Gag deletion mutants: identification of the p10 domain as a morphological determinant in the formation of spherical particles.J Virol. 1997 Jun;71(6):4425-35. doi: 10.1128/JVI.71.6.4425-4435.1997. J Virol. 1997. PMID: 9151833 Free PMC article.
-
Dimeric rous sarcoma virus capsid protein structure relevant to immature Gag assembly.J Mol Biol. 2004 Jan 2;335(1):275-82. doi: 10.1016/j.jmb.2003.10.034. J Mol Biol. 2004. PMID: 14659756
-
Virus maturation by budding.Microbiol Mol Biol Rev. 1998 Dec;62(4):1171-90. doi: 10.1128/MMBR.62.4.1171-1190.1998. Microbiol Mol Biol Rev. 1998. PMID: 9841669 Free PMC article. Review.
Cited by
-
Cooperative effect of gag proteins p12 and capsid during early events of murine leukemia virus replication.J Virol. 2005 Apr;79(7):4159-69. doi: 10.1128/JVI.79.7.4159-4169.2005. J Virol. 2005. PMID: 15767417 Free PMC article.
-
The noncanonical Gag domains p8 and n are critical for assembly and release of mouse mammary tumor virus.J Virol. 2010 Nov;84(21):11555-9. doi: 10.1128/JVI.00652-10. Epub 2010 Aug 25. J Virol. 2010. PMID: 20739518 Free PMC article.
-
Mutations affecting cleavage at the p10-capsid protease cleavage site block Rous sarcoma virus replication.Retrovirology. 2005 Sep 27;2:58. doi: 10.1186/1742-4690-2-58. Retrovirology. 2005. PMID: 16188035 Free PMC article.
-
Assembly and architecture of HIV.Adv Exp Med Biol. 2012;726:441-65. doi: 10.1007/978-1-4614-0980-9_20. Adv Exp Med Biol. 2012. PMID: 22297526 Free PMC article. Review.
-
The retroviral capsid domain dictates virion size, morphology, and coassembly of gag into virus-like particles.J Virol. 2005 Nov;79(21):13463-72. doi: 10.1128/JVI.79.21.13463-13472.2005. J Virol. 2005. PMID: 16227267 Free PMC article.
References
-
- Ausubel F M, et al., editors. Current protocols in molecular biology. New York, N.Y: John Wiley & Sons, Inc.; 1994.
-
- Bancroft J B. The self-assembly of spherical plant viruses. Adv Virus Res. 1970;16:99–134. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
