

Bim1p/Yeb1p mediates the Kar9p-dependent cortical attachment of cytoplasmic microtubules
- PMID: 10982392
- PMCID: PMC14967
- DOI: 10.1091/mbc.11.9.2949
Bim1p/Yeb1p mediates the Kar9p-dependent cortical attachment of cytoplasmic microtubules
Abstract
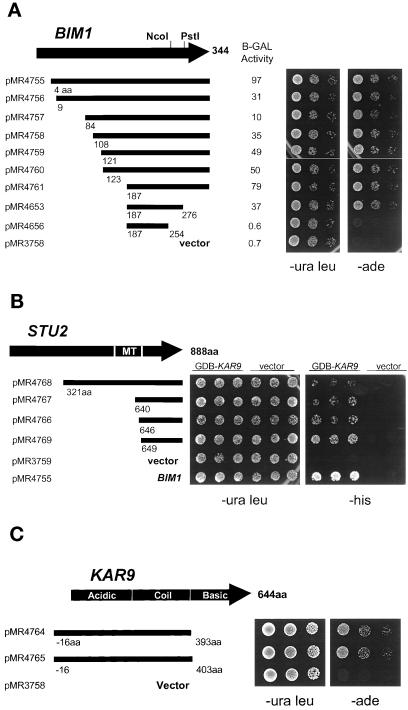
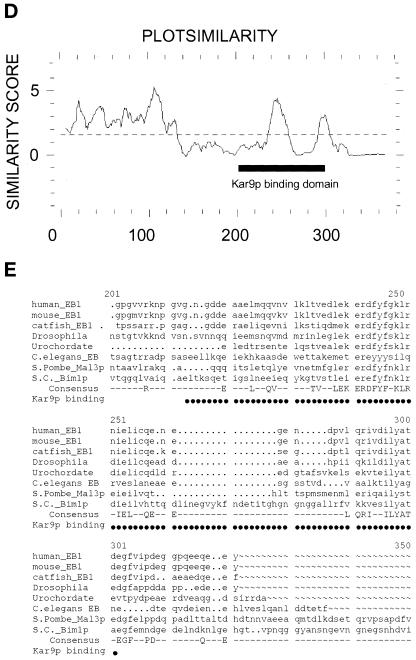
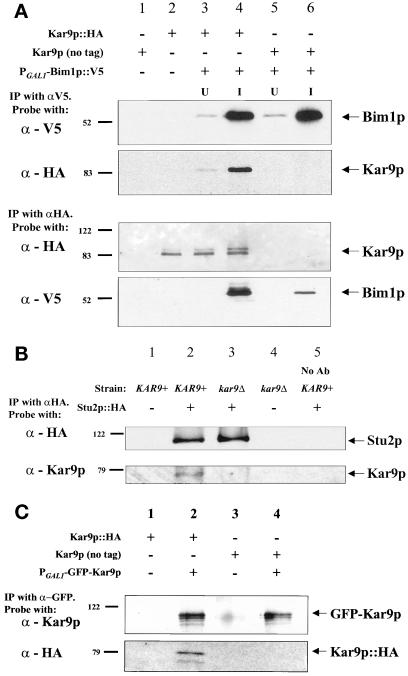
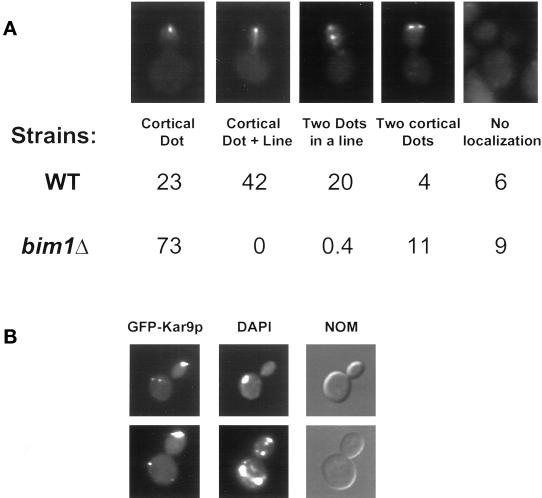
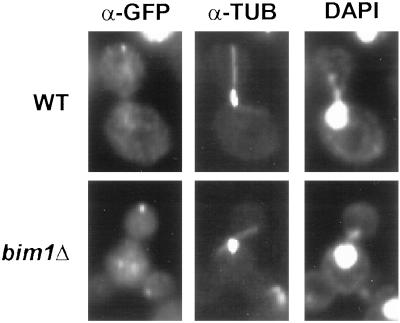
In Saccharomyces cerevisiae, positioning of the mitotic spindle depends on the interaction of cytoplasmic microtubules with the cell cortex. In this process, cortical Kar9p in the bud acts as a link between the actin and microtubule cytoskeletons. To identify Kar9p-interacting proteins, a two-hybrid screen was conducted with the use of full-length Kar9p as bait, and three genes were identified: BIM1, STU2, and KAR9 itself. STU2 encodes a component of the spindle pole body. Bim1p is the yeast homologue of the human microtubule-binding protein EB1, which is a binding partner to the adenomatous polyposis coli protein involved in colon cancer. Eighty-nine amino acids within the third quarter of Bim1p was sufficient to confer interaction with Kar9p. The two-hybrid interactions were confirmed with the use of coimmunoprecipitation experiments. Genetic analysis placed Bim1p in the Kar9p pathway for nuclear migration. Bim1p was not required for Kar9p's cortical or spindle pole body localization. However, deletion of BIM1 eliminated Kar9p localization along cytoplasmic microtubules. Furthermore, in the bim1 mutants, the cytoplasmic microtubules no longer intersected the cortical dot of Green Fluorescent Protein-Kar9p. These experiments demonstrate that the interaction of cytoplasmic microtubules with the Kar9p cortical attachment site requires the microtubule-binding protein Bim1p.
Figures
Similar articles
-
The cortical localization of the microtubule orientation protein, Kar9p, is dependent upon actin and proteins required for polarization.J Cell Biol. 1999 Mar 8;144(5):963-75. doi: 10.1083/jcb.144.5.963. J Cell Biol. 1999. PMID: 10085294 Free PMC article.
-
Positioning of the mitotic spindle by a cortical-microtubule capture mechanism.Science. 2000 Mar 24;287(5461):2260-2. doi: 10.1126/science.287.5461.2260. Science. 2000. PMID: 10731147
-
BIM1 encodes a microtubule-binding protein in yeast.Mol Biol Cell. 1997 Dec;8(12):2677-91. doi: 10.1091/mbc.8.12.2677. Mol Biol Cell. 1997. PMID: 9398684 Free PMC article.
-
Search, capture and signal: games microtubules and centrosomes play.J Cell Sci. 2001 Jan;114(Pt 2):247-55. doi: 10.1242/jcs.114.2.247. J Cell Sci. 2001. PMID: 11148127 Review.
-
Microtubule cytoskeleton: navigating the intracellular landscape.Curr Biol. 2003 May 27;13(11):R430-2. doi: 10.1016/s0960-9822(03)00362-2. Curr Biol. 2003. PMID: 12781150 Review.
Cited by
-
Mitotic spindle form and function.Genetics. 2012 Apr;190(4):1197-224. doi: 10.1534/genetics.111.128710. Genetics. 2012. PMID: 22491889 Free PMC article.
-
Stu2p, the budding yeast member of the conserved Dis1/XMAP215 family of microtubule-associated proteins is a plus end-binding microtubule destabilizer.J Cell Biol. 2003 Apr 28;161(2):359-69. doi: 10.1083/jcb.200211097. J Cell Biol. 2003. PMID: 12719475 Free PMC article.
-
Gamma-tubulin is required for proper recruitment and assembly of Kar9-Bim1 complexes in budding yeast.Mol Biol Cell. 2006 Oct;17(10):4420-34. doi: 10.1091/mbc.e06-03-0245. Epub 2006 Aug 9. Mol Biol Cell. 2006. PMID: 16899509 Free PMC article.
-
The XMAP215-family protein DdCP224 is required for cortical interactions of microtubules.BMC Cell Biol. 2004 Jun 8;5:24. doi: 10.1186/1471-2121-5-24. BMC Cell Biol. 2004. PMID: 15186508 Free PMC article.
-
The microtubule plus-end binding protein EB1 functions in root responses to touch and gravity signals in Arabidopsis.Plant Cell. 2008 Feb;20(2):396-410. doi: 10.1105/tpc.107.056846. Epub 2008 Feb 15. Plant Cell. 2008. PMID: 18281505 Free PMC article.
References
-
- Barker N, Morin PJ, Clevers H. The yin-yang of TCF/beta-catenin signaling. Adv Cancer Res. 2000;77:1–24. - PubMed
-
- Barth AI, Nathke IS, Nelson WJ. Cadherins, catenins and APC protein: interplay between cytoskeletal complexes and signaling pathways. Curr Opin Cell Biol. 1997;9:683–690. - PubMed
-
- Byers B. Cytology of the yeast life cycle. In: Strathern J, Jones EW, Broach JR, editors. The Molecular Biology of the Yeast Saccharomyces: Life Cycle and Inheritance. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory; 1981. pp. 59–96.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
