

Cotranslational dimerization of the Rel homology domain of NF-kappaB1 generates p50-p105 heterodimers and is required for effective p50 production
- PMID: 10970863
- PMCID: PMC302078
- DOI: 10.1093/emboj/19.17.4712
Cotranslational dimerization of the Rel homology domain of NF-kappaB1 generates p50-p105 heterodimers and is required for effective p50 production
Abstract
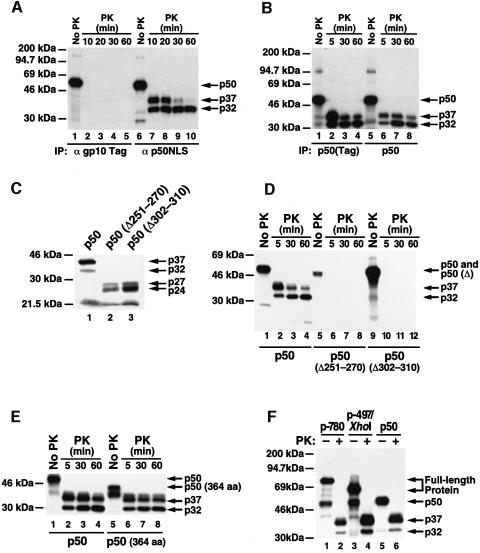
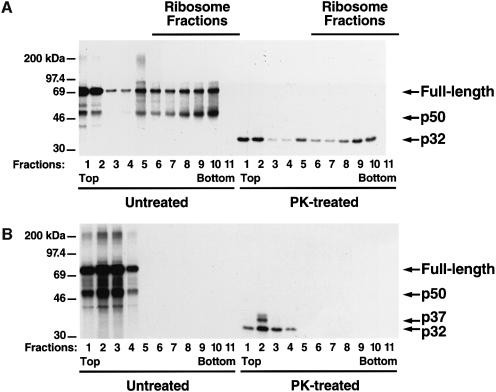
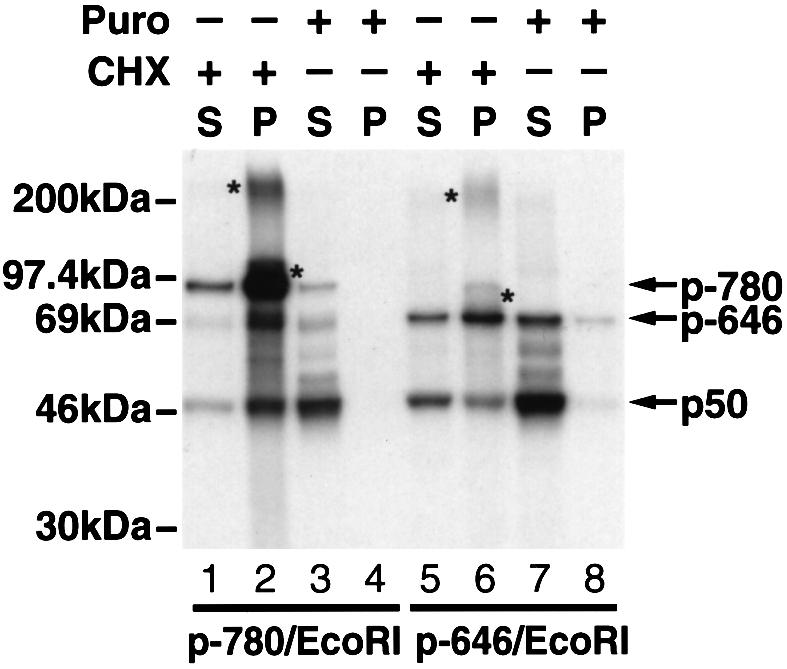
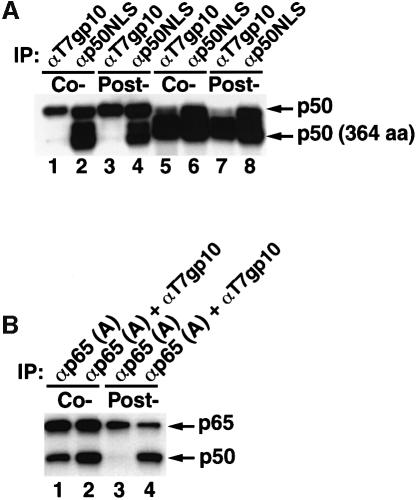
Generation of the NF-kappaB p50 transcription factor is mediated by the proteasome. We found previously that p50 is generated during translation of the NFKB1 gene and that this cotranslational processing allows the production of both p50 and p105 from a single mRNA. We now demonstrate that the Rel homology domain in p50 undergoes cotranslational dimerization and that this interaction is required for efficient production of p50. We further show that this coupling of dimerization and proteasome processing during translation uniquely generates p50-p105 heterodimers. Accordingly, after the primary cotranslational event, additional posttranslational steps regulate p50 homodimer formation and the intracellular ratio of p50 and p105. This cellular strategy places p50 under the control of the p105 inhibitor early in its biogenesis, thereby regulating the pool of p50 homodimers within the cell.
Figures

Similar articles
-
TPL-2 kinase regulates the proteolysis of the NF-kappaB-inhibitory protein NF-kappaB1 p105.Nature. 1999 Jan 28;397(6717):363-8. doi: 10.1038/16946. Nature. 1999. PMID: 9950430
-
The activity of a 70 kilodalton I kappa B molecule identical to the carboxyl terminus of the p105 NF-kappa B precursor is modulated by protein kinase A.Cell Growth Differ. 1993 Aug;4(8):617-27. Cell Growth Differ. 1993. PMID: 8398903
-
Inhibition of transcription by B cell Leukemia 3 (Bcl-3) protein requires interaction with nuclear factor κB (NF-κB) p50.J Biol Chem. 2014 Mar 7;289(10):7059-7067. doi: 10.1074/jbc.M114.551986. Epub 2014 Jan 23. J Biol Chem. 2014. PMID: 24459141 Free PMC article.
-
Nuclear factor-kappaB1: regulation and function.Int J Biochem Cell Biol. 2008;40(8):1425-30. doi: 10.1016/j.biocel.2007.05.004. Epub 2007 May 17. Int J Biochem Cell Biol. 2008. PMID: 17693123 Review.
-
The NF-ĸB p50 subunit generated by KPC1-mediated ubiquitination and limited proteasomal processing, suppresses tumor growth.Cancer Cell Int. 2023 Apr 13;23(1):67. doi: 10.1186/s12935-023-02919-5. Cancer Cell Int. 2023. PMID: 37055826 Free PMC article. Review.
Cited by
-
The mammalian cholesterol synthesis enzyme squalene monooxygenase is proteasomally truncated to a constitutively active form.J Biol Chem. 2021 Jan-Jun;296:100731. doi: 10.1016/j.jbc.2021.100731. Epub 2021 Apr 30. J Biol Chem. 2021. PMID: 33933449 Free PMC article.
-
N-glycosylation enhances presentation of a MHC class I-restricted epitope from tyrosinase.J Immunol. 2009 Apr 15;182(8):4830-5. doi: 10.4049/jimmunol.0802902. J Immunol. 2009. PMID: 19342661 Free PMC article.
-
Polyribosomes are molecular 3D nanoprinters that orchestrate the assembly of vault particles.ACS Nano. 2014 Nov 25;8(11):11552-9. doi: 10.1021/nn504778h. Epub 2014 Oct 30. ACS Nano. 2014. PMID: 25354757 Free PMC article.
-
Loss of parasympathetic innervation leads to sustained expression of pro-inflammatory genes in the rat lacrimal gland.Auton Neurosci. 2006 Jan 30;124(1-2):81-9. doi: 10.1016/j.autneu.2005.12.001. Epub 2006 Feb 2. Auton Neurosci. 2006. PMID: 16458615 Free PMC article.
-
Exploring major signaling cascades in melanomagenesis: a rationale route for targetted skin cancer therapy.Biosci Rep. 2018 Oct 2;38(5):BSR20180511. doi: 10.1042/BSR20180511. Print 2018 Oct 31. Biosci Rep. 2018. PMID: 30166456 Free PMC article. Review.
References
-
- Baeuerle P.A. and Baltimore,D. (1988) IκB: a specific inhibitor of the NF-κB transcription factor. Science, 242, 540–546. - PubMed
-
- Baeuerle P.A. and Baltimore,D. (1989) A 65-kD subunit of active NF-κB is required for inhibition of NF–κB by IκB. Genes Dev., 3, 1689–1698. - PubMed
-
- Baumeister W., Walz,J., Zühl,F. and Seemüller,E. (1998) The proteasome: paradigm of a self-compartmentalizing protease. Cell, 92, 367–380. - PubMed
-
- Belich M.P., Salmerón,A., Johnson,L.H. and Ley,S.C. (1999) TPL-2 kinase regulates the proteolysis of the NF-κB-inhibitory protein NF-κB 1 p105. Nature, 397, 363–368. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
