

Genetic interactions between TFIIS and the Swi-Snf chromatin-remodeling complex
- PMID: 10913179
- PMCID: PMC86073
- DOI: 10.1128/MCB.20.16.5960-5973.2000
Genetic interactions between TFIIS and the Swi-Snf chromatin-remodeling complex
Abstract
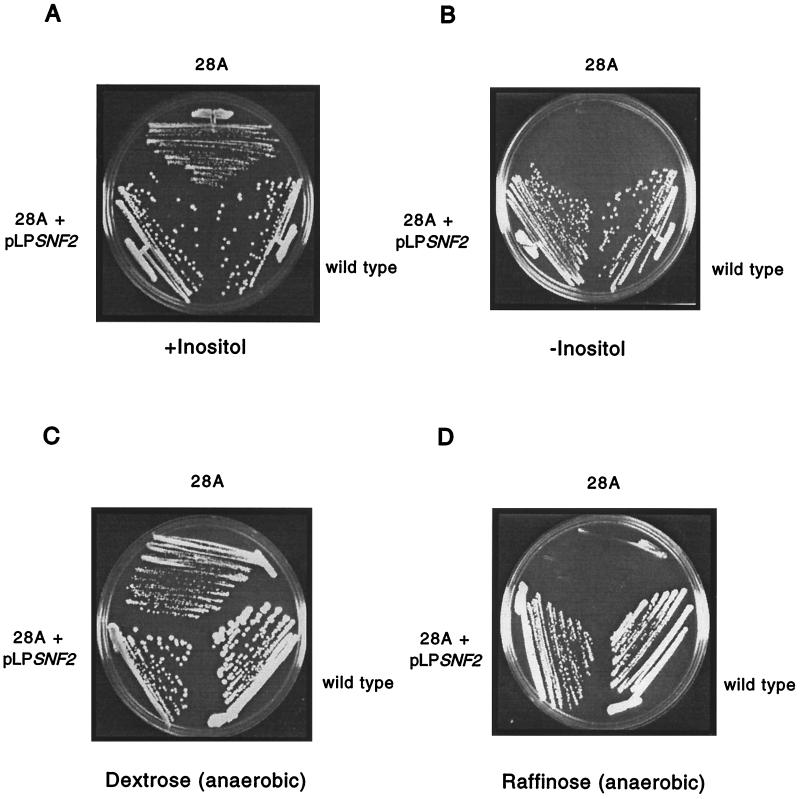
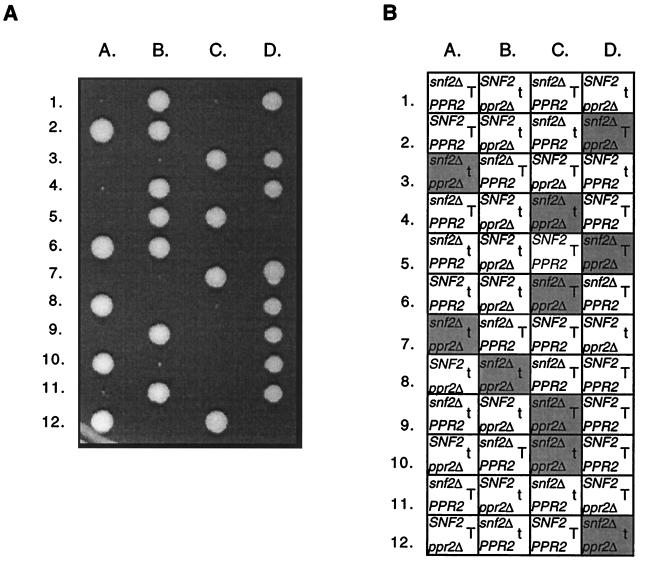
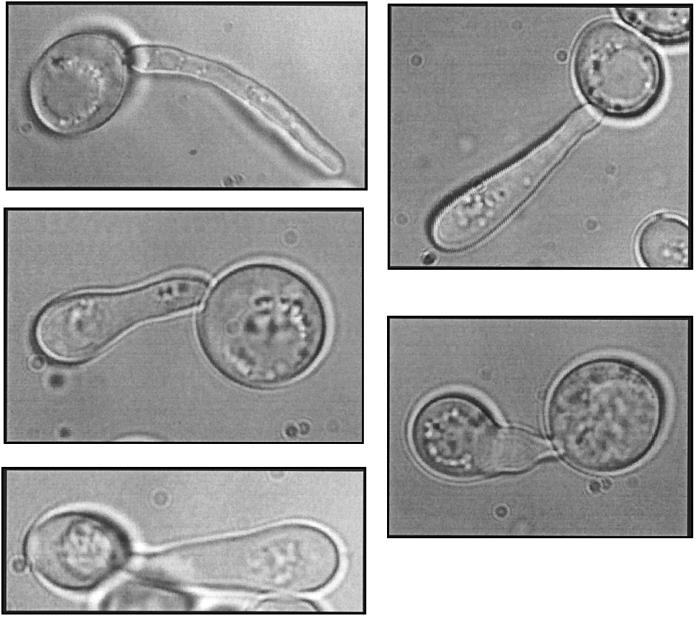
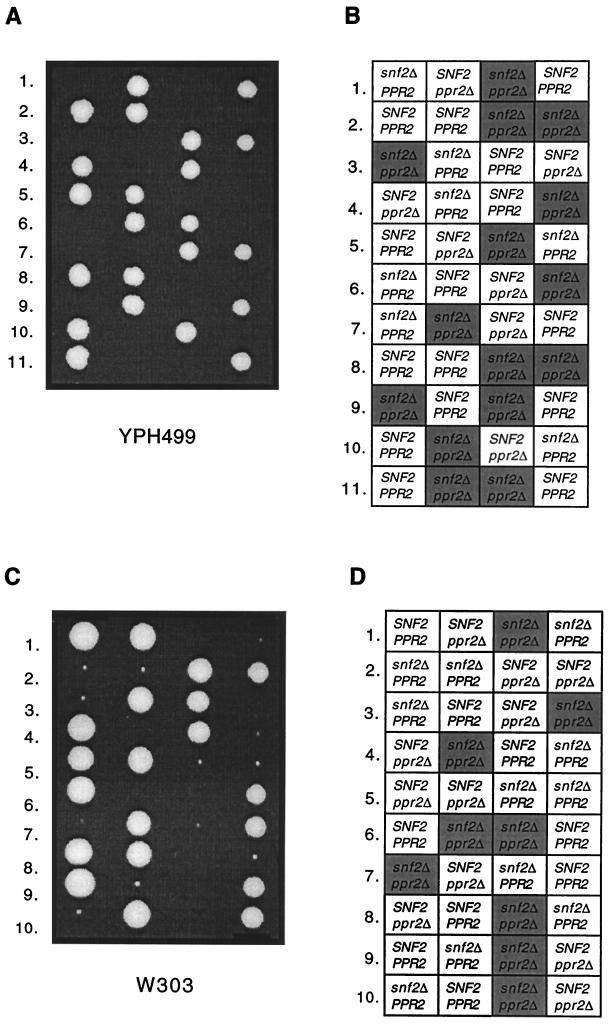
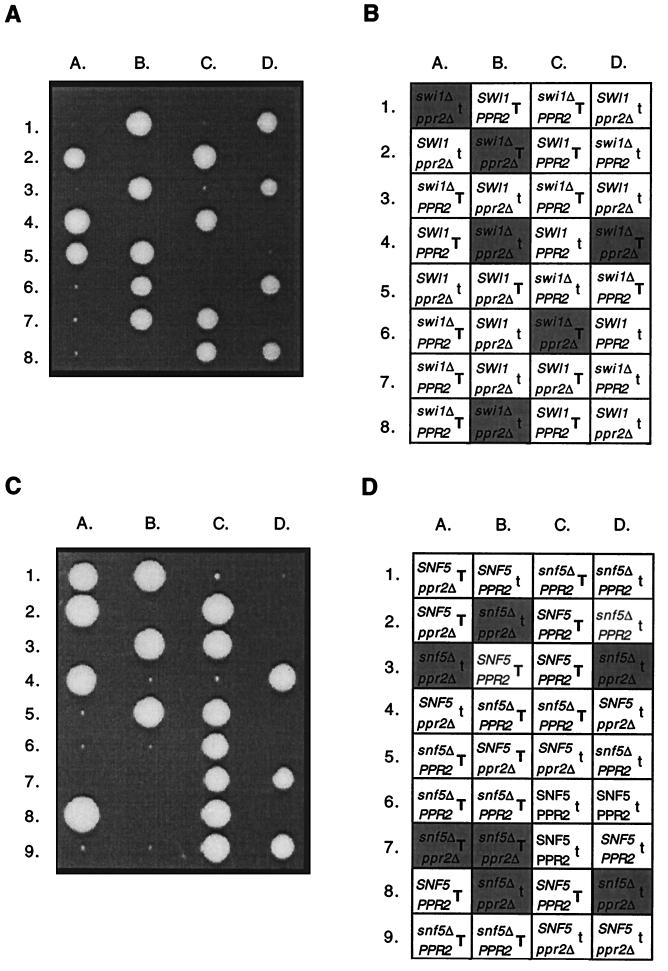
The eukaryotic transcript elongation factor TFIIS enables RNA polymerase II to read through blocks to elongation in vitro and interacts genetically with a variety of components of the transcription machinery in vivo. In Saccharomyces cerevisiae, the gene encoding TFIIS (PPR2) is not essential, and disruption strains exhibit only mild phenotypes and an increased sensitivity to 6-azauracil. The nonessential nature of TFIIS encouraged the use of a synthetic lethal screen to elucidate the in vivo roles of TFIIS as well as provide more information on other factors involved in the regulation of transcript elongation. Several genes were identified that are necessary for either cell survival or robust growth when the gene encoding TFIIS has been disrupted. These include UBP3, KEX2, STT4, and SWI2/SNF2. SWI1 and SNF5 disruptions were also synthetically lethal with ppr2Delta, suggesting that the reduced ability to remodel chromatin confers the synthetic phenotype. The synthetic phenotypes show marked osmosensitivity and cytoskeletal defects, including a terminal hyperelongated bud phenotype with the Swi-Snf complex. These results suggest that genes important in osmoregulation, cell membrane synthesis and integrity, and cell division may require the Swi-Snf complex and TFIIS for efficient transcription. The detection of these genetic interactions provides another functional link between the Swi-Snf complex and the elongation machinery.
Figures
Similar articles
-
Genetic interactions between TFIIF and TFIIS.Genetics. 2006 Aug;173(4):1871-84. doi: 10.1534/genetics.106.058834. Epub 2006 Apr 30. Genetics. 2006. PMID: 16648643 Free PMC article.
-
Subunits of the yeast SWI/SNF complex are members of the actin-related protein (ARP) family.J Biol Chem. 1998 Sep 11;273(37):23641-4. doi: 10.1074/jbc.273.37.23641. J Biol Chem. 1998. PMID: 9726966
-
Interaction of a Swi3 homolog with Sth1 provides evidence for a Swi/Snf-related complex with an essential function in Saccharomyces cerevisiae.Mol Cell Biol. 1997 Apr;17(4):1768-75. doi: 10.1128/MCB.17.4.1768. Mol Cell Biol. 1997. PMID: 9121424 Free PMC article.
-
The mammalian SWI/SNF complex and the control of cell growth.Semin Cell Dev Biol. 1999 Apr;10(2):189-95. doi: 10.1006/scdb.1999.0300. Semin Cell Dev Biol. 1999. PMID: 10441072 Review.
-
Promoter targeting and chromatin remodeling by the SWI/SNF complex.Curr Opin Genet Dev. 2000 Apr;10(2):187-92. doi: 10.1016/s0959-437x(00)00068-x. Curr Opin Genet Dev. 2000. PMID: 10753786 Review.
Cited by
-
Analysis of gene induction and arrest site transcription in yeast with mutations in the transcription elongation machinery.J Biol Chem. 2001 Apr 13;276(15):11531-8. doi: 10.1074/jbc.M011322200. Epub 2001 Jan 19. J Biol Chem. 2001. PMID: 11278887 Free PMC article.
-
The multifunctional Ccr4-Not complex directly promotes transcription elongation.Genes Dev. 2011 Mar 15;25(6):581-93. doi: 10.1101/gad.2020911. Genes Dev. 2011. PMID: 21406554 Free PMC article.
-
Members of the SAGA and Mediator complexes are partners of the transcription elongation factor TFIIS.EMBO J. 2004 Oct 27;23(21):4232-42. doi: 10.1038/sj.emboj.7600326. Epub 2004 Sep 9. EMBO J. 2004. PMID: 15359273 Free PMC article.
-
Use of an in vivo reporter assay to test for transcriptional and translational fidelity in yeast.J Biol Chem. 2002 Jul 5;277(27):24420-6. doi: 10.1074/jbc.M202059200. Epub 2002 May 2. J Biol Chem. 2002. PMID: 12006589 Free PMC article.
-
The basal initiation machinery: beyond the general transcription factors.Curr Opin Cell Biol. 2009 Jun;21(3):344-51. doi: 10.1016/j.ceb.2009.03.006. Epub 2009 May 4. Curr Opin Cell Biol. 2009. PMID: 19411170 Free PMC article. Review.
References
-
- Boeke J D, Trueheart J, Natsoulis G, Fink G R. 5-Fluoroorotic acid as a selective agent in yeast molecular genetics. Methods Enzymol. 1987;154:164–175. - PubMed
-
- Bortvin A, Winston F. Evidence that Spt6p controls chromatin structure by a direct interaction with histones. Science. 1996;272:1473–1476. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases