

Cloning and mutagenesis of the murine gammaherpesvirus 68 genome as an infectious bacterial artificial chromosome
- PMID: 10888635
- PMCID: PMC112213
- DOI: 10.1128/jvi.74.15.6964-6974.2000
Cloning and mutagenesis of the murine gammaherpesvirus 68 genome as an infectious bacterial artificial chromosome
Abstract
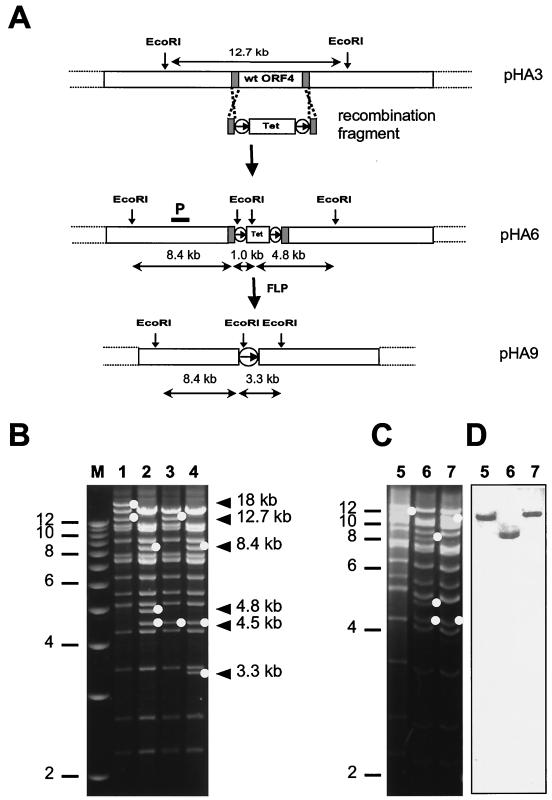
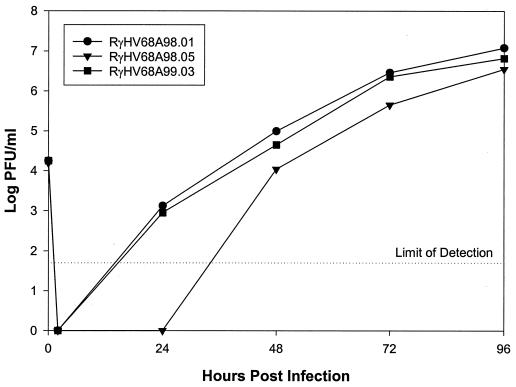
Gammaherpesviruses cause important infections of humans, in particular in immunocompromised patients. Recently, murine gammaherpesvirus 68 (MHV-68) infection of mice has been developed as a small animal model of gammaherpesvirus pathogenesis. Efficient generation of mutants of MHV-68 would significantly contribute to the understanding of viral gene functions in virus-host interaction, thereby further enhancing the potential of this model. To this end, we cloned the MHV-68 genome as a bacterial artificial chromosome (BAC) in Escherichia coli. During propagation in E. coli, spontaneous recombination events within the internal and terminal repeats of the cloned MHV-68 genome, affecting the copy number of the repeats, were occasionally observed. The gene for the green fluorescent protein was incorporated into the cloned BAC for identification of infected cells. BAC vector sequences were flanked by loxP sites to allow the excision of these sequences using recombinase Cre and to allow the generation of recombinant viruses with wild-type genome properties. Infectious virus was reconstituted from the BAC-cloned MHV-68. Growth of the BAC-derived virus in cell culture was indistinguishable from that of wild-type MHV-68. To assess the feasibility of mutagenesis of the cloned MHV-68 genome, a mutant virus with a deletion of open reading frame 4 was generated. Genetically modified MHV-68 can now be analyzed in functionally modified mouse strains to assess the role of gammaherpesvirus genes in virus-host interaction and pathogenesis.
Figures




Similar articles
-
Virus reconstituted from infectious bacterial artificial chromosome (BAC)-cloned murine gammaherpesvirus 68 acquires wild-type properties in vivo only after excision of BAC vector sequences.J Virol. 2001 Jun;75(12):5692-6. doi: 10.1128/JVI.75.12.5692-5696.2001. J Virol. 2001. PMID: 11356978 Free PMC article.
-
Construction and characterization of an infectious murine gammaherpesivrus-68 bacterial artificial chromosome.J Biomed Biotechnol. 2011;2011:926258. doi: 10.1155/2011/926258. Epub 2010 Dec 9. J Biomed Biotechnol. 2011. PMID: 21197474 Free PMC article.
-
Cloning of herpesviral genomes as bacterial artificial chromosomes.Rev Med Virol. 2003 Mar-Apr;13(2):111-21. doi: 10.1002/rmv.380. Rev Med Virol. 2003. PMID: 12627394 Review.
-
Construction of a self-excisable bacterial artificial chromosome containing the human cytomegalovirus genome and mutagenesis of the diploid TRL/IRL13 gene.J Virol. 2002 Mar;76(5):2316-28. doi: 10.1128/jvi.76.5.2316-2328.2002. J Virol. 2002. PMID: 11836410 Free PMC article.
-
Biological and pathogenetic characterization of different isolates of murine gammaherpesvirus 68 (MHV-68) in the context of study of human oncogenic gammaherpesviruses.Acta Virol. 2013;57(2):105-12. doi: 10.4149/av_2013_02_105. Acta Virol. 2013. PMID: 23600868 Review.
Cited by
-
A replication-deficient gammaherpesvirus vaccine protects mice from lytic disease and reduces latency establishment.NPJ Vaccines. 2024 Jun 24;9(1):116. doi: 10.1038/s41541-024-00908-x. NPJ Vaccines. 2024. PMID: 38914546 Free PMC article.
-
Gammaherpesvirus infection triggers the formation of tRNA fragments from premature tRNAs.bioRxiv [Preprint]. 2024 May 2:2024.05.01.592122. doi: 10.1101/2024.05.01.592122. bioRxiv. 2024. PMID: 38746336 Free PMC article. Preprint.
-
Gammaherpesvirus infection alters transfer RNA splicing and triggers tRNA cleavage.bioRxiv [Preprint]. 2024 Feb 17:2024.02.16.580780. doi: 10.1101/2024.02.16.580780. bioRxiv. 2024. PMID: 38405876 Free PMC article. Preprint.
-
A simple method for rapid cloning of complete herpesvirus genomes.Cell Rep Methods. 2024 Feb 26;4(2):100696. doi: 10.1016/j.crmeth.2024.100696. Epub 2024 Jan 23. Cell Rep Methods. 2024. PMID: 38266652 Free PMC article.
-
A Mouse Model to Study the Pathogenesis of γ-herpesviral Infections in Germinal Center B Cells.Cells. 2023 Dec 6;12(24):2780. doi: 10.3390/cells12242780. Cells. 2023. PMID: 38132100 Free PMC article.
References
-
- Blaskovic D, Stancekova M, Svobodova J, Mistrikova J. Isolation of five strains of herpesviruses from two species of free living small rodents. Acta Virol. 1980;24:468. - PubMed
-
- Brune W, Menard C, Hobom U, Odenbreit S, Messerle M, Koszinowski U H. Rapid identification of essential and nonessential herpesvirus genes by direct transposon mutagenesis. Nat Biotechnol. 1999;17:360–364. - PubMed
-
- Cherepanov P P, Wackernagel W. Gene disruption in Escherichia coli: TcR and KmR cassettes with the option of Flp-catalyzed excision of the antibiotic-resistance determinant. Gene. 1995;158:9–14. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
