

Identification of a novel family of nonclassic yeast phosphatidylinositol transfer proteins whose function modulates phospholipase D activity and Sec14p-independent cell growth
- PMID: 10848624
- PMCID: PMC14898
- DOI: 10.1091/mbc.11.6.1989
Identification of a novel family of nonclassic yeast phosphatidylinositol transfer proteins whose function modulates phospholipase D activity and Sec14p-independent cell growth
Abstract
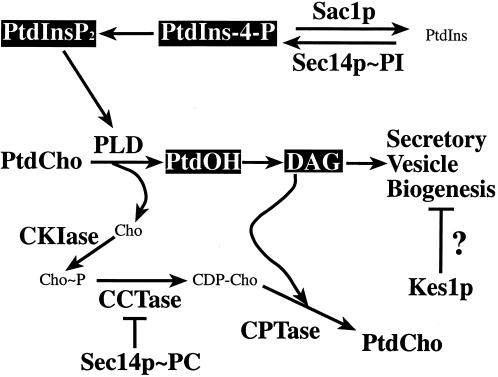
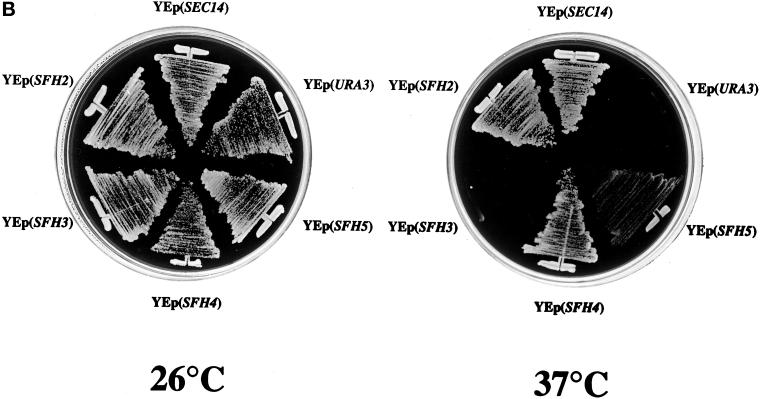
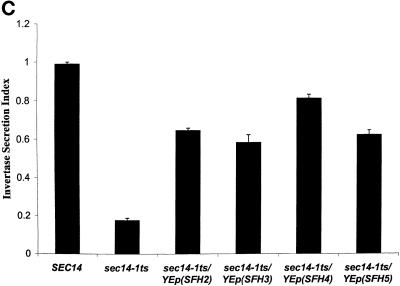
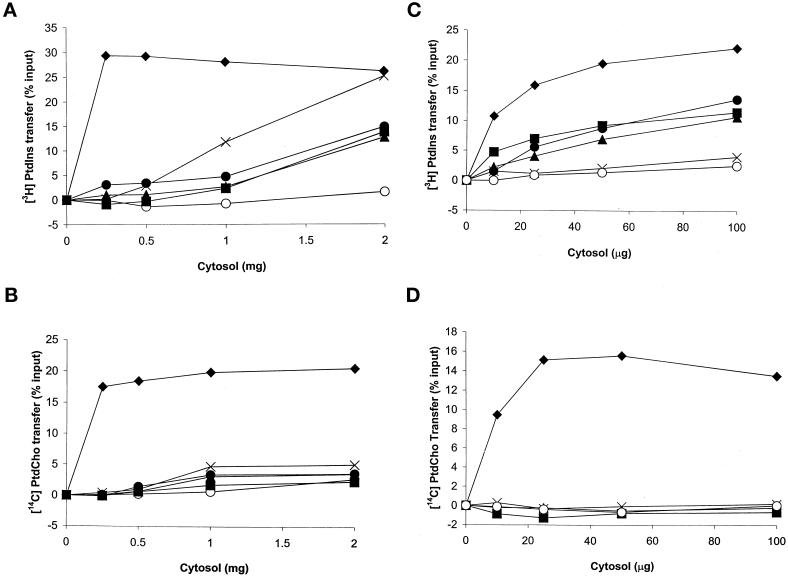
Yeast phosphatidylinositol transfer protein (Sec14p) is essential for Golgi function and cell viability. We now report a characterization of five yeast SFH (Sec Fourteen Homologue) proteins that share 24-65% primary sequence identity with Sec14p. We show that Sfh1p, which shares 64% primary sequence identity with Sec14p, is nonfunctional as a Sec14p in vivo or in vitro. Yet, SFH proteins sharing low primary sequence similarity with Sec14p (i.e., Sfh2p, Sfh3p, Sfh4p, and Sfh5p) represent novel phosphatidylinositol transfer proteins (PITPs) that exhibit phosphatidylinositol- but not phosphatidylcholine-transfer activity in vitro. Moreover, increased expression of Sfh2p, Sfh4p, or Sfh5p rescues sec14-associated growth and secretory defects in a phospholipase D (PLD)-sensitive manner. Several independent lines of evidence further demonstrate that SFH PITPs are collectively required for efficient activation of PLD in vegetative cells. These include a collective requirement for SFH proteins in Sec14p-independent cell growth and in optimal activation of PLD in Sec14p-deficient cells. Consistent with these findings, Sfh2p colocalizes with PLD in endosomal compartments. The data indicate that SFH gene products cooperate with "bypass-Sec14p" mutations and PLD in a complex interaction through which yeast can adapt to loss of the essential function of Sec14p. These findings expand the physiological repertoire of PITP function in yeast and provide the first in vivo demonstration of a role for specific PITPs in stimulating activation of PLD.
Figures

Similar articles
-
Subcellular localization of yeast Sec14 homologues and their involvement in regulation of phospholipid turnover.Eur J Biochem. 2003 Aug;270(15):3133-45. doi: 10.1046/j.1432-1033.2003.03688.x. Eur J Biochem. 2003. PMID: 12869188
-
Phospholipase D activity is required for suppression of yeast phosphatidylinositol transfer protein defects.Proc Natl Acad Sci U S A. 1998 Oct 13;95(21):12346-51. doi: 10.1073/pnas.95.21.12346. Proc Natl Acad Sci U S A. 1998. PMID: 9770489 Free PMC article.
-
Nonclassical PITPs activate PLD via the Stt4p PtdIns-4-kinase and modulate function of late stages of exocytosis in vegetative yeast.Traffic. 2005 Dec;6(12):1157-72. doi: 10.1111/j.1600-0854.2005.00350.x. Traffic. 2005. PMID: 16262726
-
Sec14p-like proteins regulate phosphoinositide homoeostasis and intracellular protein and lipid trafficking in yeast.Biochem Soc Trans. 2006 Jun;34(Pt 3):346-50. doi: 10.1042/BST0340346. Biochem Soc Trans. 2006. PMID: 16709158 Review.
-
Sec14 related proteins in yeast.Biochim Biophys Acta. 2007 Jun;1771(6):737-45. doi: 10.1016/j.bbalip.2007.02.008. Epub 2007 Mar 2. Biochim Biophys Acta. 2007. PMID: 17395532 Review.
Cited by
-
Sec14-nodulin proteins and the patterning of phosphoinositide landmarks for developmental control of membrane morphogenesis.Mol Biol Cell. 2015 May 1;26(9):1764-81. doi: 10.1091/mbc.E14-10-1475. Epub 2015 Mar 4. Mol Biol Cell. 2015. PMID: 25739452 Free PMC article.
-
Phosphatidylcholine synthesis influences the diacylglycerol homeostasis required for SEC14p-dependent Golgi function and cell growth.Mol Biol Cell. 2001 Mar;12(3):511-20. doi: 10.1091/mbc.12.3.511. Mol Biol Cell. 2001. PMID: 11251067 Free PMC article.
-
Functional diversification of the chemical landscapes of yeast Sec14-like phosphatidylinositol transfer protein lipid-binding cavities.J Biol Chem. 2019 Dec 13;294(50):19081-19098. doi: 10.1074/jbc.RA119.011153. Epub 2019 Nov 5. J Biol Chem. 2019. PMID: 31690622 Free PMC article.
-
Genomewide Elucidation of Drug Resistance Mechanisms for Systemically Used Antifungal Drugs Amphotericin B, Caspofungin, and Voriconazole in the Budding Yeast.Antimicrob Agents Chemother. 2019 Aug 23;63(9):e02268-18. doi: 10.1128/AAC.02268-18. Print 2019 Sep. Antimicrob Agents Chemother. 2019. PMID: 31209012 Free PMC article.
-
A blueprint for functional engineering: Single point mutations reconstitute phosphatidylinositol presentation in a pseudo-Sec14 protein.Commun Integr Biol. 2011 Nov 1;4(6):674-8. doi: 10.4161/cib.17064. Commun Integr Biol. 2011. PMID: 22446525 Free PMC article.
References
-
- Aitken JF, van Heusden GPH, Temkin M, Dowhan W. The gene encoding the phosphatidylinositol transfer protein is essential for cell growth. J Biol Chem. 1990;265:4711–4717. - PubMed
-
- Bankaitis VA, Aitken JR, Cleves AE, Dowhan W. An essential role for a phospholipid transfer protein in yeast Golgi function. Nature. 1990;347:561–562. - PubMed
-
- Bankaitis VA, Fry MR, Cartee RT, Kagiwada S. Molecular Biology Intelligence Unit. Austin, TX: RG Landes; 1996. Phospholipid transfer proteins: emerging roles in vesicle trafficking, signal transduction, and metabolic regulation; pp. 94–97.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
