

Evolution and assembly of an extremely scrambled gene
- PMID: 10725348
- PMCID: PMC16233
- DOI: 10.1073/pnas.97.7.3298
Evolution and assembly of an extremely scrambled gene
Abstract
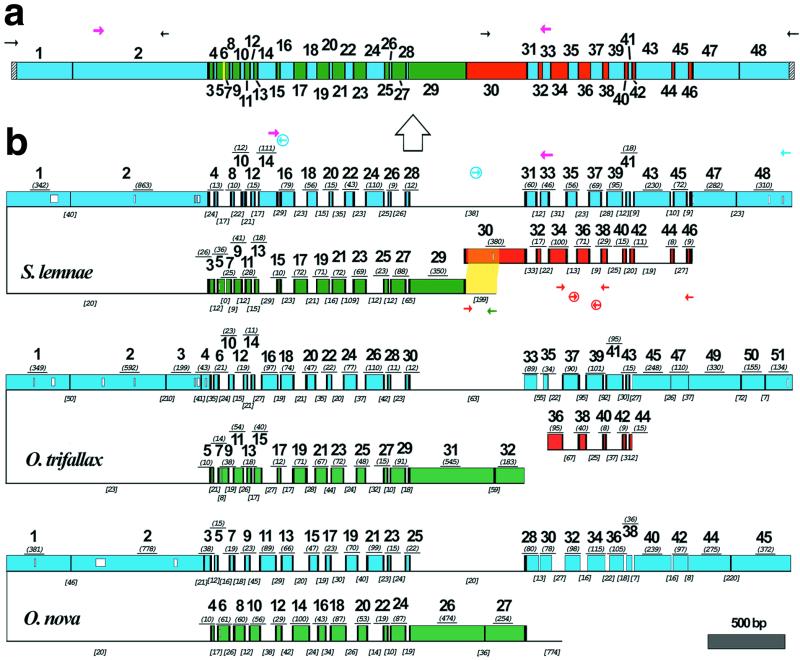
The process of gene unscrambling in hypotrichous ciliates represents one of nature's ingenious solutions to the problem of gene assembly. With some essential genes scrambled in as many as 51 pieces, these ciliates rely on sequence and structural cues to rebuild their fragmented genes and genomes. Here we report the complex pattern of scrambling in the DNA polymerase alpha gene of Stylonychia lemnae. The germline (micronuclear) copy of this gene is broken into 48 pieces with 47 dispersed over two loci, with no asymmetry in the placement of coding segments on either strand. Direct repeats present at the boundaries between coding and noncoding sequences provide pointers to help guide assembly of the functional (macronuclear) gene. We investigate the evolution of this complex gene in three hypotrichous species.
Figures


Similar articles
-
The evolutionary scrambling and developmental unscrambling of germline genes in hypotrichous ciliates.Nucleic Acids Res. 1999 Mar 1;27(5):1243-50. doi: 10.1093/nar/27.5.1243. Nucleic Acids Res. 1999. PMID: 9973610 Free PMC article. Review.
-
Evolution of internal eliminated segments and scrambling in the micronuclear gene encoding DNA polymerase alpha in two Oxytricha species.Nucleic Acids Res. 1997 May 15;25(10):1883-9. doi: 10.1093/nar/25.10.1883. Nucleic Acids Res. 1997. PMID: 9115353 Free PMC article.
-
The germline gene encoding DNA polymerase alpha in the hypotrichous ciliate Oxytricha nova is extremely scrambled.Nucleic Acids Res. 1996 Sep 1;24(17):3337-40. doi: 10.1093/nar/24.17.3337. Nucleic Acids Res. 1996. PMID: 8811087 Free PMC article.
-
Polymorphism, recombination and alternative unscrambling in the DNA polymerase alpha gene of the ciliate Stylonychia lemnae (Alveolata; class Spirotrichea).Genetics. 2003 Dec;165(4):1761-77. doi: 10.1093/genetics/165.4.1761. Genetics. 2003. PMID: 14704164 Free PMC article.
-
Internal eliminated segments (IESs) of Oxytrichidae.J Eukaryot Microbiol. 1996 Nov-Dec;43(6):432-41. doi: 10.1111/j.1550-7408.1996.tb04502.x. J Eukaryot Microbiol. 1996. PMID: 8976601 Review.
Cited by
-
Analyses of alternatively processed genes in ciliates provide insights into the origins of scrambled genomes and may provide a mechanism for speciation.mBio. 2015 Feb 3;6(1):e01998-14. doi: 10.1128/mBio.01998-14. mBio. 2015. PMID: 25650397 Free PMC article.
-
Interconversion of germline-limited and somatic DNA in a scrambled gene.J Mol Evol. 2006 Jul;63(1):69-73. doi: 10.1007/s00239-005-0166-4. Epub 2006 Jun 3. J Mol Evol. 2006. PMID: 16755354
-
Programmed genome rearrangements in ciliates.Cell Mol Life Sci. 2020 Nov;77(22):4615-4629. doi: 10.1007/s00018-020-03555-2. Epub 2020 May 27. Cell Mol Life Sci. 2020. PMID: 32462406 Free PMC article. Review.
-
RNA-Mediated Epigenetic Programming of Genome Rearrangements.Annu Rev Genomics Hum Genet. 2011;12:367-89. doi: 10.1146/annurev-genom-082410-101420. Annu Rev Genomics Hum Genet. 2011. PMID: 21801022 Free PMC article. Review.
-
Intron Evolution and Information processing in the DNA polymerase alpha gene in spirotrichous ciliates: a hypothesis for interconversion between DNA and RNA deletion.Biol Direct. 2007 Feb 1;2:6. doi: 10.1186/1745-6150-2-6. Biol Direct. 2007. PMID: 17270054 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
