

Alpha/beta interferon protects adult mice from fatal Sindbis virus infection and is an important determinant of cell and tissue tropism
- PMID: 10708454
- PMCID: PMC111838
- DOI: 10.1128/jvi.74.7.3366-3378.2000
Alpha/beta interferon protects adult mice from fatal Sindbis virus infection and is an important determinant of cell and tissue tropism
Abstract
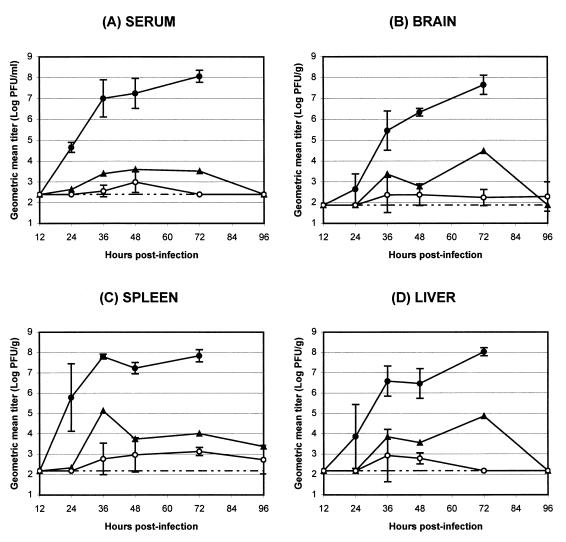
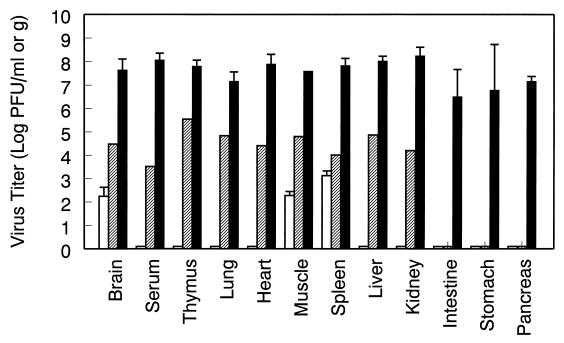
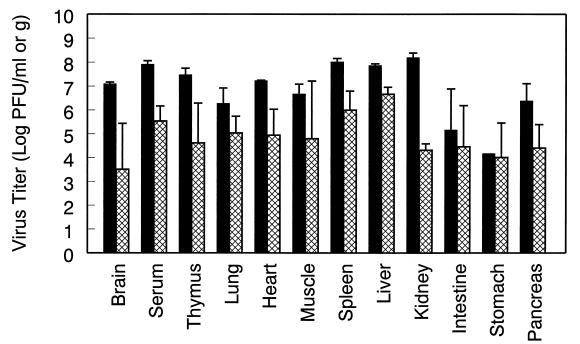
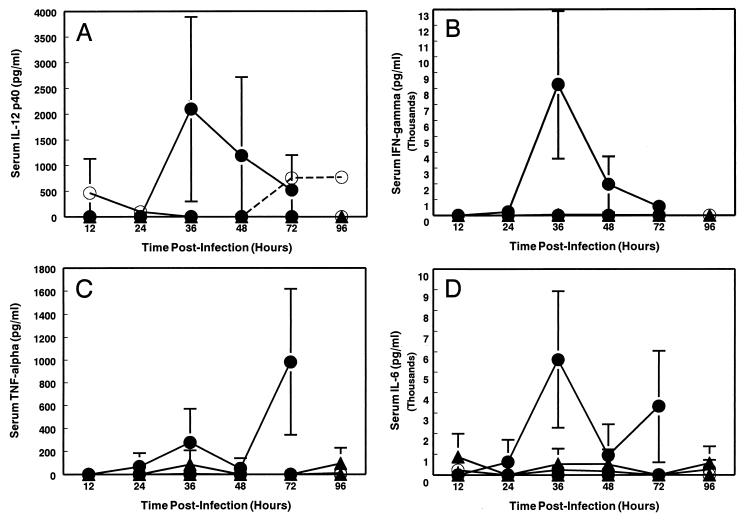
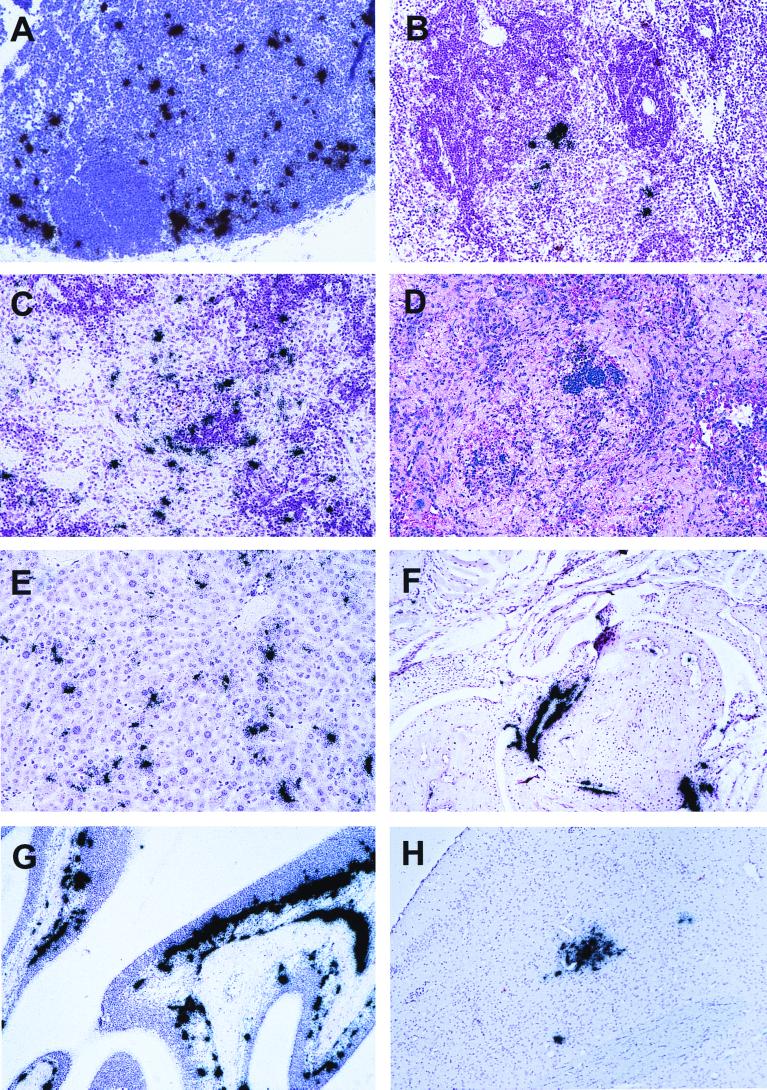
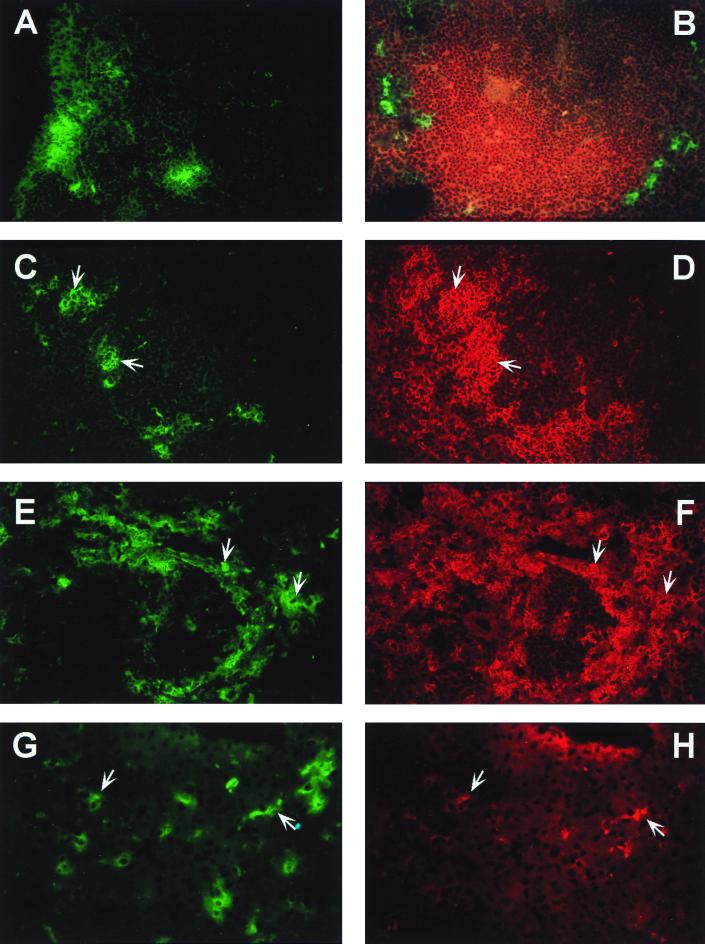
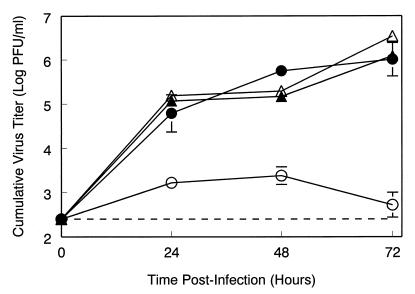
Infection of adult 129 Sv/Ev mice with consensus Sindbis virus strain TR339 is subclinical due to an inherent restriction in early virus replication and viremic dissemination. By comparing the pathogenesis of TR339 in 129 Sv/Ev mice and alpha/beta interferon receptor null (IFN-alpha/betaR(-/-)) mice, we have assessed the contribution of IFN-alpha/beta in restricting virus replication and spread and in determining cell and tissue tropism. In adult 129 Sv/Ev mice, subcutaneous inoculation with 100 PFU of TR339 led to extremely low-level virus replication and viremia, with clearance under way by 96 h postinoculation (p.i.). In striking contrast, adult IFN-alpha/betaR(-/-) mice inoculated subcutaneously with 100 PFU of TR339 succumbed to the infection within 84 h. By 24 h p.i. a high-titer serum viremia had seeded infectious virus systemically, coincident with the systemic induction of the proinflammatory cytokines interleukin-12 (IL-12) p40, IFN-gamma, tumor necrosis factor alpha, and IL-6. Replicating virus was located in macrophage-dendritic cell (DC)-like cells at 24 h p.i. in the draining lymph node and in the splenic marginal zone. By 72 h p.i. virus replication was widespread in macrophage-DC-like cells in the spleen, liver, lung, thymus, and kidney and in fibroblast-connective tissue and periosteum, with sporadic neuroinvasion. IFN-alpha/beta-mediated restriction of TR339 infection was mimicked in vitro in peritoneal exudate cells from 129 Sv/Ev versus IFN-alpha/betaR(-/-) mice. Thus, IFN-alpha/beta protects the normal adult host from viral infection by rapidly conferring an antiviral state on otherwise permissive cell types, both locally and systemically. Ablation of the IFN-alpha/beta system alters the apparent cell and tissue tropism of the virus and renders macrophage-DC-lineage cells permissive to infection.
Figures
Similar articles
-
Effects of PKR/RNase L-dependent and alternative antiviral pathways on alphavirus replication and pathogenesis.Viral Immunol. 2002;15(1):53-76. doi: 10.1089/088282402317340233. Viral Immunol. 2002. PMID: 11952147
-
Roles of nonstructural protein nsP2 and Alpha/Beta interferons in determining the outcome of Sindbis virus infection.J Virol. 2002 Nov;76(22):11254-64. doi: 10.1128/jvi.76.22.11254-11264.2002. J Virol. 2002. PMID: 12388685 Free PMC article.
-
Infection of neonatal mice with sindbis virus results in a systemic inflammatory response syndrome.J Virol. 1999 Dec;73(12):10387-98. doi: 10.1128/JVI.73.12.10387-10398.1999. J Virol. 1999. PMID: 10559357 Free PMC article.
-
The role of antibody in recovery from alphavirus encephalitis.Immunol Rev. 1997 Oct;159:155-61. doi: 10.1111/j.1600-065x.1997.tb01013.x. Immunol Rev. 1997. PMID: 9416509 Review.
-
Age-dependent susceptibility to fatal encephalitis: alphavirus infection of neurons.Arch Virol Suppl. 1994;9:31-9. doi: 10.1007/978-3-7091-9326-6_4. Arch Virol Suppl. 1994. PMID: 8032263 Review.
Cited by
-
Mouse macrophage innate immune response to Chikungunya virus infection.Virol J. 2012 Dec 19;9:313. doi: 10.1186/1743-422X-9-313. Virol J. 2012. PMID: 23253140 Free PMC article.
-
Myd88-dependent toll-like receptor 7 signaling mediates protection from severe Ross River virus-induced disease in mice.J Virol. 2012 Oct;86(19):10675-85. doi: 10.1128/JVI.00601-12. Epub 2012 Jul 25. J Virol. 2012. PMID: 22837203 Free PMC article.
-
Gene expression in the brain during reovirus encephalitis.J Neurovirol. 2010 Feb;16(1):56-71. doi: 10.3109/13550280903586394. J Neurovirol. 2010. PMID: 20158406 Free PMC article.
-
Gamma interferon-dependent, noncytolytic clearance of sindbis virus infection from neurons in vitro.J Virol. 2005 May;79(9):5374-85. doi: 10.1128/JVI.79.9.5374-5385.2005. J Virol. 2005. PMID: 15827152 Free PMC article.
-
Naturally Occurring and Engineered Alphaviruses Sensitive to Double-Stranded-RNA-Activated Protein Kinase Show Restricted Translation in Mammalian Cells, Increased Sensitivity to Interferon, and Marked Oncotropism.J Virol. 2020 Jan 17;94(3):e01630-19. doi: 10.1128/JVI.01630-19. Print 2020 Jan 17. J Virol. 2020. PMID: 31723025 Free PMC article.
References
-
- Agger R, Crowley M T, Witmer-Pack M D. The surface of dendritic cells in the mouse as studied with monoclonal antibodies. Int Rev Immunol. 1990;6:89–101. - PubMed
-
- Billiau A, Vandekerckhove F. Cytokines and their interactions with other inflammatory mediators in the pathogenesis of sepsis and septic shock. Eur J Clin Investig. 1991;21:559–573. - PubMed
-
- Biron C A. Initial and innate responses to viral infections—pattern setting in immunity or disease. Curr Opin Microbiol. 1999;2:374–381. - PubMed
-
- Boonpucknavig S, Vuttiviroj O, Boonpucknavig V. Infection of young adult mice with dengue virus type 2. Trans R Soc Trop Med Hyg. 1981;75:647–653. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
