

bcl-2 transgene expression inhibits apoptosis in the germinal center and reveals differences in the selection of memory B cells and bone marrow antibody-forming cells
- PMID: 10662793
- PMCID: PMC2195819
- DOI: 10.1084/jem.191.3.475
bcl-2 transgene expression inhibits apoptosis in the germinal center and reveals differences in the selection of memory B cells and bone marrow antibody-forming cells
Abstract
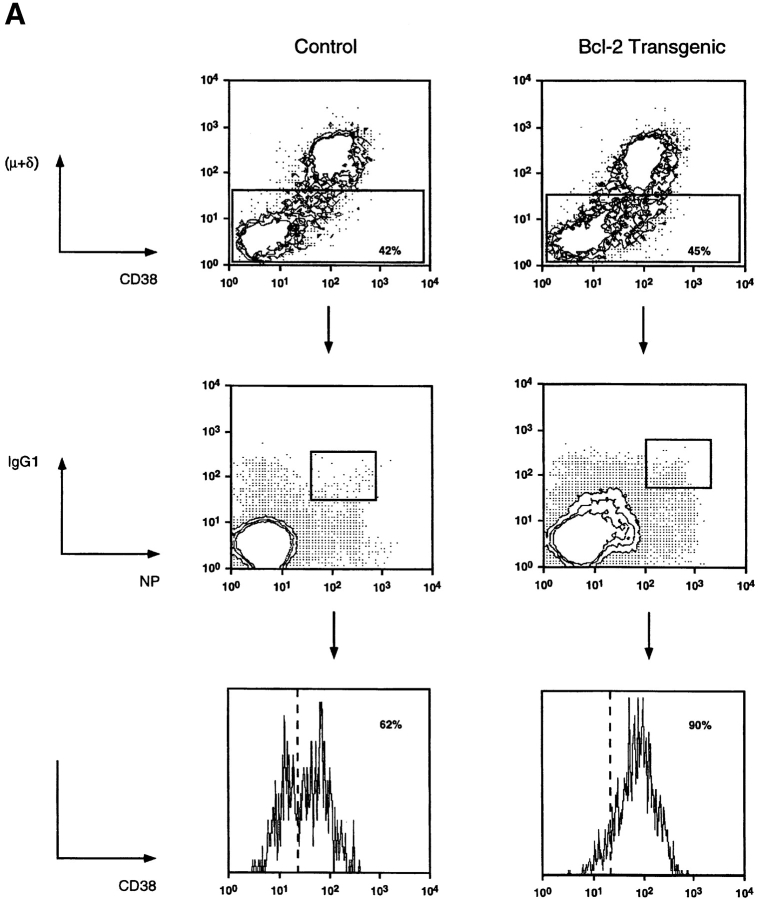
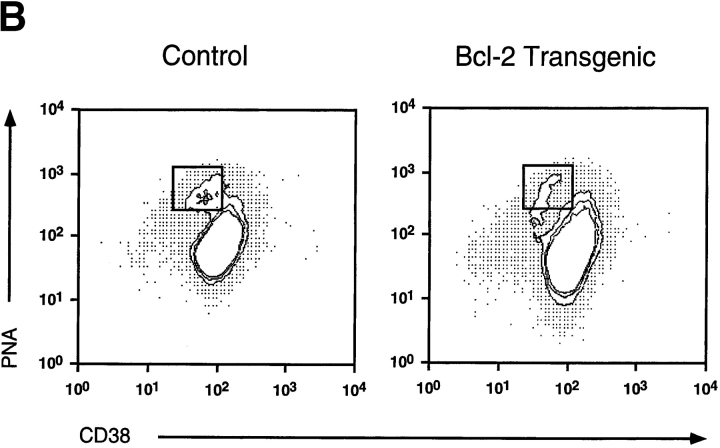
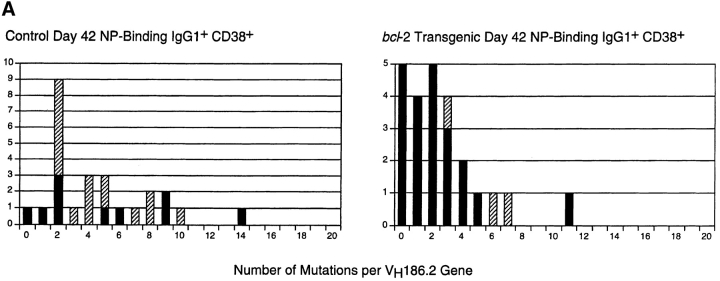
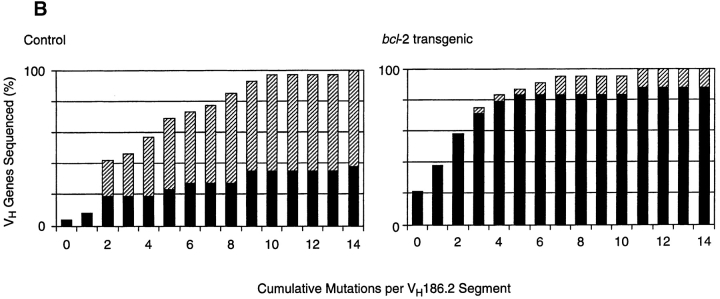
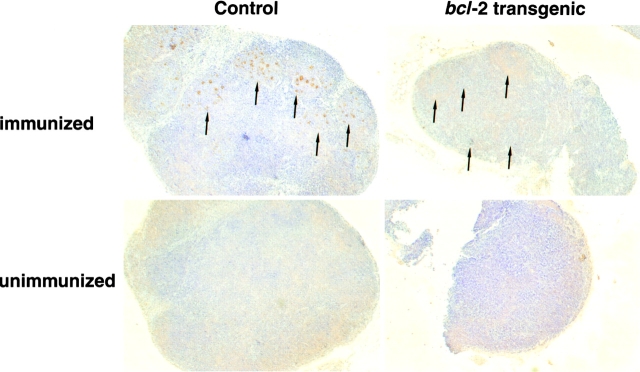
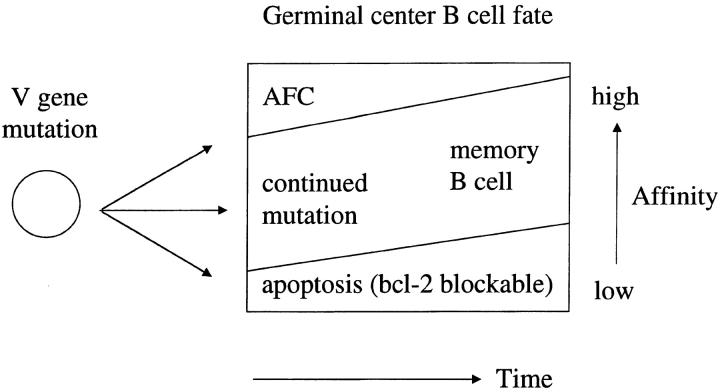
Immunization with T cell-dependent antigens generates long-lived memory B cells and antibody-forming cells (AFCs). Both populations originate in germinal centers and, predominantly, produce antibodies with high affinity for antigen. The means by which germinal center B cells are recruited into these populations remains unclear. We have examined affinity maturation of antigen-specific B cells in mice expressing the cell death inhibitor bcl-2 as a transgene. Such mice had reduced apoptosis in germinal centers and an excessive number of memory B cells with a low frequency of V gene somatic mutation, including those mutations encoding amino acid exchanges known to enhance affinity. Despite the frequency of AFCs being increased in bcl-2-transgenic mice, the fraction secreting high-affinity antibody in the bone marrow at day 42 remained unchanged compared with controls. The inability of BCL-2 to alter selection of bone marrow AFCs is consistent with these cells being selected within the germinal center on the basis of their affinity being above some threshold rather than their survival being due to a selective competition for an antigen-based signal. Continuous competition for antigen does, however, explain formation of the memory compartment.
Figures
Similar articles
-
Relaxed negative selection in germinal centers and impaired affinity maturation in bcl-xL transgenic mice.J Exp Med. 1999 Aug 2;190(3):399-410. doi: 10.1084/jem.190.3.399. J Exp Med. 1999. PMID: 10430628 Free PMC article.
-
Germinal center founder cells display propensity for apoptosis before onset of somatic mutation.J Exp Med. 1997 Feb 3;185(3):563-71. doi: 10.1084/jem.185.3.563. J Exp Med. 1997. PMID: 9053456 Free PMC article.
-
Kinetics of establishing the memory B cell population as revealed by CD38 expression.J Immunol. 1998 May 15;160(10):4688-95. J Immunol. 1998. PMID: 9590214
-
B lymphocyte physiology: the beginning and the end.Ciba Found Symp. 1997;204:220-30; discussion 230-1. doi: 10.1002/9780470515280.ch15. Ciba Found Symp. 1997. PMID: 9107424 Review.
-
Apoptosis and the B cell response to antigen.Int Rev Immunol. 1997;15(1-2):53-71. doi: 10.3109/08830189709068171. Int Rev Immunol. 1997. PMID: 9178072 Review.
Cited by
-
The transcription factor Hhex cooperates with the corepressor Tle3 to promote memory B cell development.Nat Immunol. 2020 Sep;21(9):1082-1093. doi: 10.1038/s41590-020-0713-6. Epub 2020 Jun 29. Nat Immunol. 2020. PMID: 32601467 Free PMC article.
-
Depletion of preexisting B-cell lymphoma 2-expressing senescent cells before vaccination impacts antigen-specific antitumor immune responses in old mice.Aging Cell. 2023 Dec;22(12):e14007. doi: 10.1111/acel.14007. Epub 2023 Nov 23. Aging Cell. 2023. PMID: 37997569 Free PMC article.
-
Signals controlling rest and reactivation of T helper memory lymphocytes in bone marrow.Cell Mol Life Sci. 2012 May;69(10):1609-13. doi: 10.1007/s00018-012-0969-6. Epub 2012 Mar 30. Cell Mol Life Sci. 2012. PMID: 22460581 Free PMC article. Review.
-
Estrogen alters thresholds for B cell apoptosis and activation.J Clin Invest. 2002 Jun;109(12):1625-33. doi: 10.1172/JCI14873. J Clin Invest. 2002. PMID: 12070310 Free PMC article.
-
Germinal center B and follicular helper T cells: siblings, cousins or just good friends?Nat Immunol. 2011 Jun;12(6):472-7. doi: 10.1038/ni.2019. Nat Immunol. 2011. PMID: 21739669 Review.
References
-
- Nossal G.J. The molecular and cellular basis of affinity maturation in the antibody response. Cell. 1992;68:1–2 . - PubMed
-
- MacLennan I.C. Germinal centers. Annu. Rev. Immunol. 1994;12:117–139. - PubMed
-
- Kelsoe G. Life and death in germinal centers (redux) Immunity. 1996;4:107–111. - PubMed
-
- Weiss U., Zoebelein R., Rajewsky K. Accumulation of somatic mutants in the B cell compartment after primary immunization with a T cell-dependent antigen. Eur. J. Immunol. 1992;22:511–517. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
