

Nuclear accumulation of S-adenosylhomocysteine hydrolase in transcriptionally active cells during development of Xenopus laevis
- PMID: 10588658
- PMCID: PMC25758
- DOI: 10.1091/mbc.10.12.4283
Nuclear accumulation of S-adenosylhomocysteine hydrolase in transcriptionally active cells during development of Xenopus laevis
Abstract
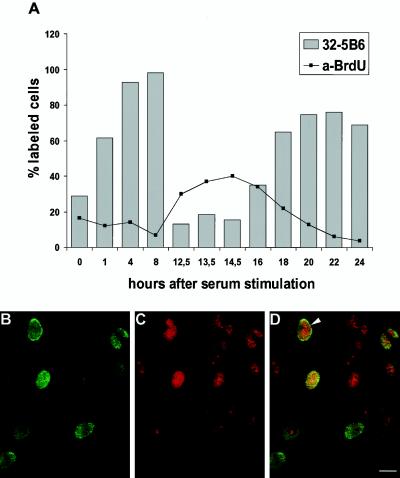
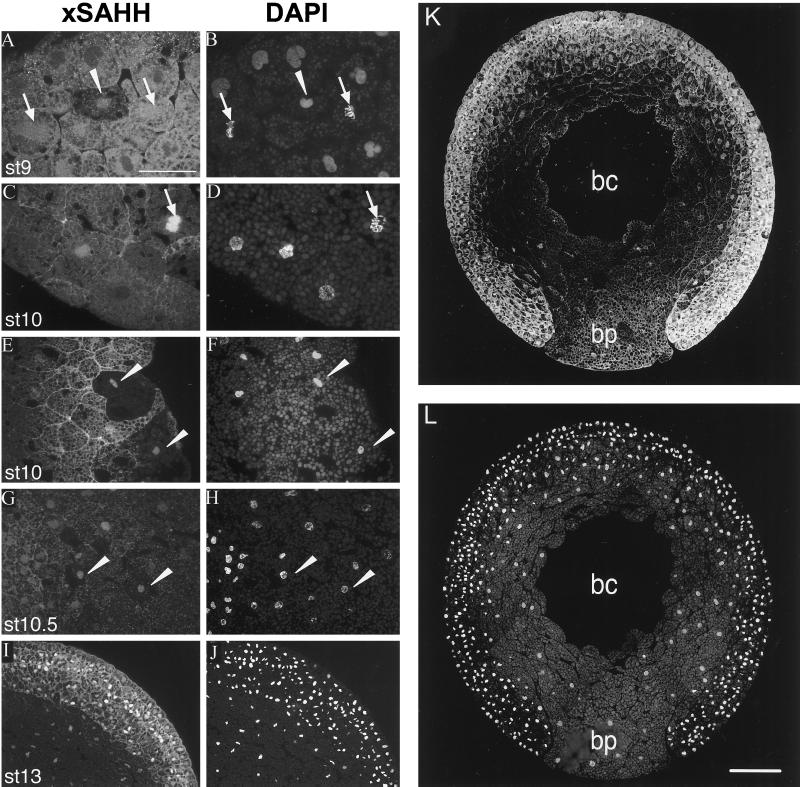
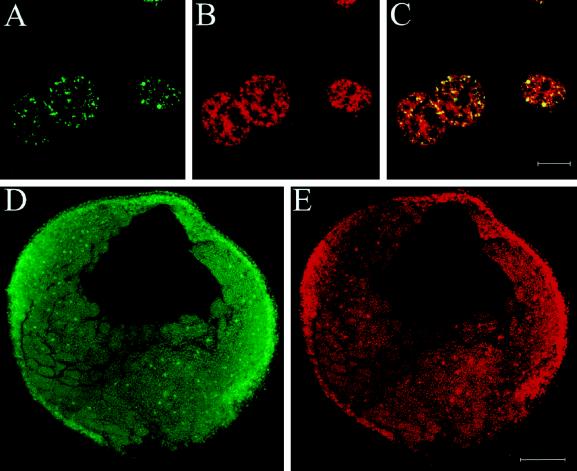
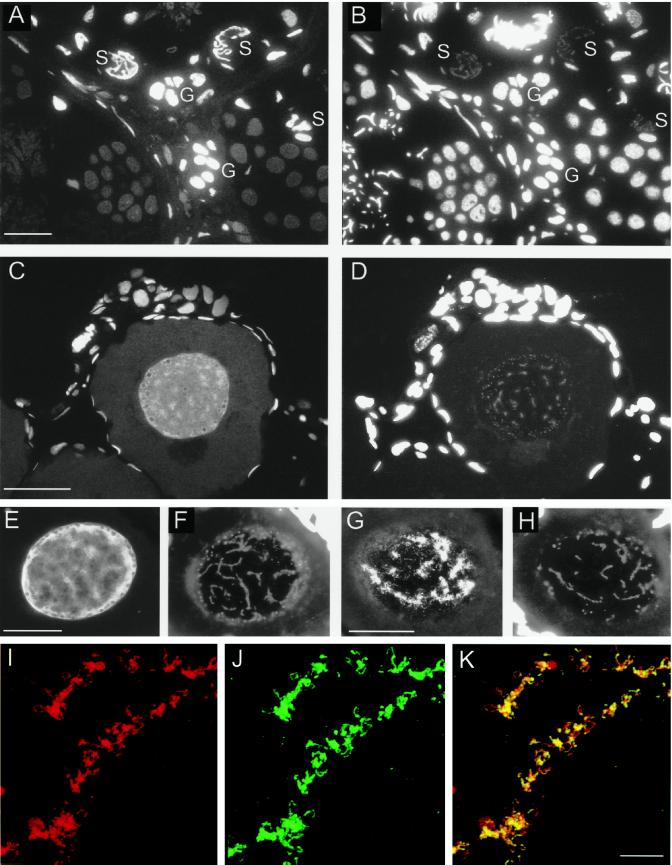
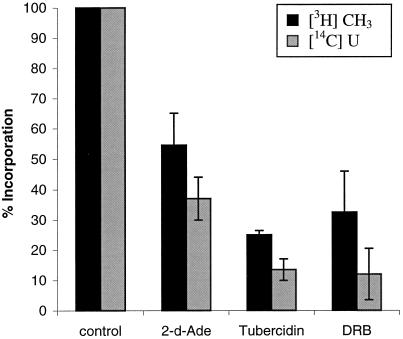
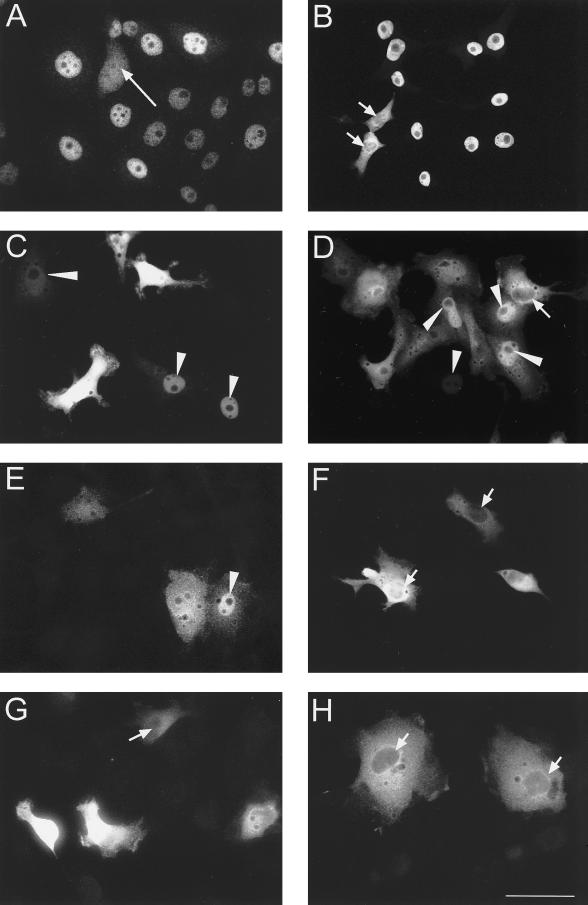
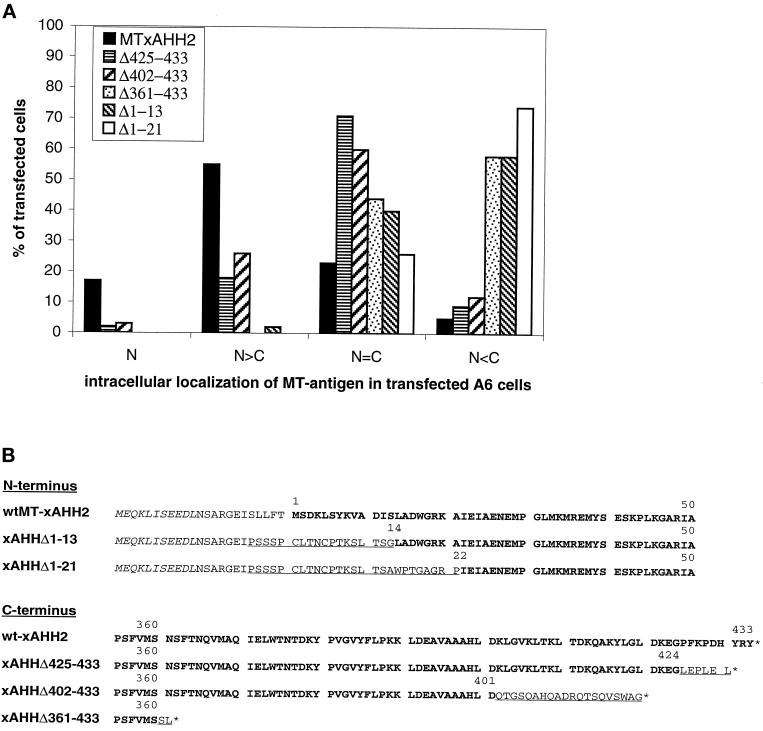
The oocyte nuclear antigen of the monoclonal antibody 32-5B6 of Xenopus laevis is subject to regulated nuclear translocation during embryogenesis. It is distributed in the cytoplasm during oocyte maturation, where it remains during cleavage and blastula stages, before it gradually reaccumulates in the nuclei during gastrulation. We have now identified this antigen to be the enzyme S-adenosylhomocysteine hydrolase (SAHH). SAHH is the only enzyme that cleaves S-adenosylhomocysteine, a reaction product and an inhibitor of all S-adenosylmethionine-dependent methylation reactions. We have compared the spatial and temporal patterns of nuclear localization of SAHH and of nuclear methyltransferase activities during embryogenesis and in tissue culture cells. Nuclear localization of Xenopus SAHH did not temporally correlate with DNA methylation. However, we found that SAHH nuclear localization coincides with high rates of mRNA synthesis, a subpopulation colocalizes with RNA polymerase II, and inhibitors of SAHH reduce both methylation and synthesis of poly(A)(+) RNA. We therefore propose that accumulation of SAHH in the nucleus may be required for efficient cap methylation in transcriptionally active cells. Mutation analysis revealed that the C terminus and the N terminus are both required for efficient nuclear translocation in tissue culture cells, indicating that more than one interacting domain contributes to nuclear accumulation of Xenopus SAHH.
Figures

Similar articles
-
Interaction of S-adenosylhomocysteine hydrolase of Xenopus laevis with mRNA(guanine-7-)methyltransferase: implication on its nuclear compartmentalisation and on cap methylation of hnRNA.Biochim Biophys Acta. 2002 Jun 12;1590(1-3):93-102. doi: 10.1016/s0167-4889(02)00205-7. Biochim Biophys Acta. 2002. PMID: 12063172
-
Nuclear targeting of methyl-recycling enzymes in Arabidopsis thaliana is mediated by specific protein interactions.Mol Plant. 2012 Jan;5(1):231-48. doi: 10.1093/mp/ssr083. Epub 2011 Oct 5. Mol Plant. 2012. PMID: 21976714
-
Poly (A) polymerases in the nucleus and cytoplasm of frog oocytes: dynamic changes during oocyte maturation and early development.RNA. 1995 Mar;1(1):64-78. RNA. 1995. PMID: 7489490 Free PMC article.
-
Requirements for nuclear translocation and nucleolar accumulation of nucleolin of Xenopus laevis.Eur J Cell Biol. 1993 Aug;61(2):369-82. Eur J Cell Biol. 1993. PMID: 8223724
-
Monoclonal antibody against dnmt1 arrests the cell division of xenopus early-stage embryos.Exp Cell Res. 2003 Jun 10;286(2):252-62. doi: 10.1016/s0014-4827(03)00060-0. Exp Cell Res. 2003. PMID: 12749854
Cited by
-
Analysis and characterization of differential gene expression during rapid trophoblastic elongation in the pig using suppression subtractive hybridization.Reprod Biol Endocrinol. 2003 Feb 14;1:23. doi: 10.1186/1477-7827-1-23. Reprod Biol Endocrinol. 2003. PMID: 12646053 Free PMC article.
-
Methylation of two-component response regulator MtrA in mycobacteria negatively modulates its DNA binding and transcriptional activation.Biochem J. 2020 Dec 11;477(23):4473-4489. doi: 10.1042/BCJ20200455. Biochem J. 2020. PMID: 33175092 Free PMC article.
-
Acute liver injury induces nucleocytoplasmic redistribution of hepatic methionine metabolism enzymes.Antioxid Redox Signal. 2014 Jun 1;20(16):2541-54. doi: 10.1089/ars.2013.5342. Epub 2014 Jan 3. Antioxid Redox Signal. 2014. PMID: 24124652 Free PMC article.
-
S-adenosyl-L-homocysteine hydrolase and methylation disorders: yeast as a model system.Biochim Biophys Acta. 2013 Jan;1832(1):204-15. doi: 10.1016/j.bbadis.2012.09.007. Epub 2012 Sep 24. Biochim Biophys Acta. 2013. PMID: 23017368 Free PMC article. Review.
-
S-adenosyl homocysteine hydrolase is required for Myc-induced mRNA cap methylation, protein synthesis, and cell proliferation.Mol Cell Biol. 2009 Dec;29(23):6182-91. doi: 10.1128/MCB.00973-09. Epub 2009 Oct 5. Mol Cell Biol. 2009. PMID: 19805518 Free PMC article.
References
-
- Ault-Riche DB, Yuan CS, Borchardt RT. A single mutation at lysine 426 of human placental S-adenosylhomocysteine hydrolase inactivates the enzyme. J Biol Chem. 1994;269:31472–31478. - PubMed
-
- Bachvarova R, Davidson EH. Nuclear activation at the onset of amphibian gastrulation. J Exp Zool. 1966;163:285–296.
-
- Bergmann S, Shatrov V, Ratter F, Schiemann S, Schulze-Osthoff K, Lehmann V. Adenosine and homocysteine together enhance TNF-mediated cytotoxicity but do not alter activation of nuclear factor-kappa B in L929 cells. J Immunol. 1994;153:1736–1743. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
