

Promiscuity in Rab-SNARE interactions
- PMID: 10588649
- PMCID: PMC25749
- DOI: 10.1091/mbc.10.12.4149
Promiscuity in Rab-SNARE interactions
Abstract
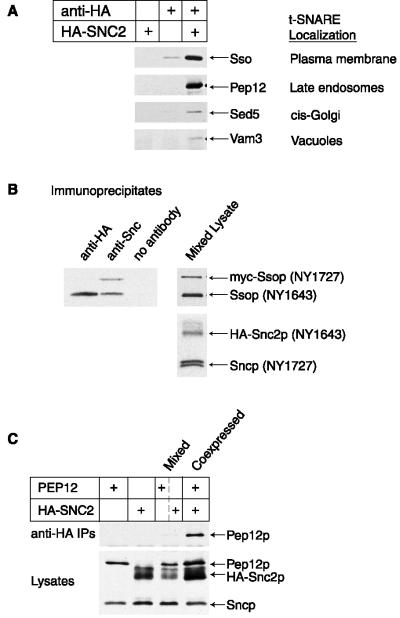
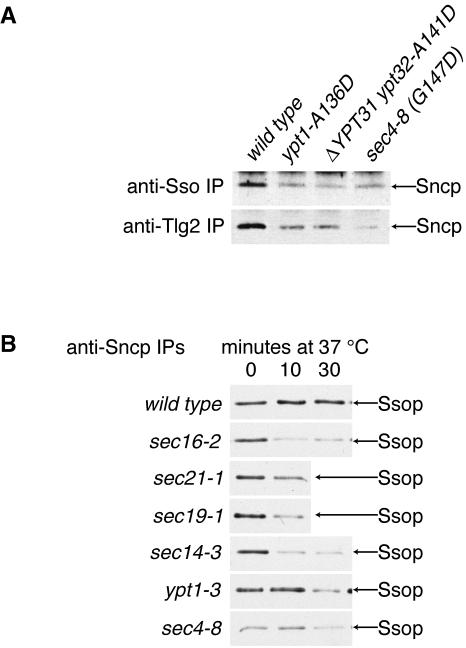
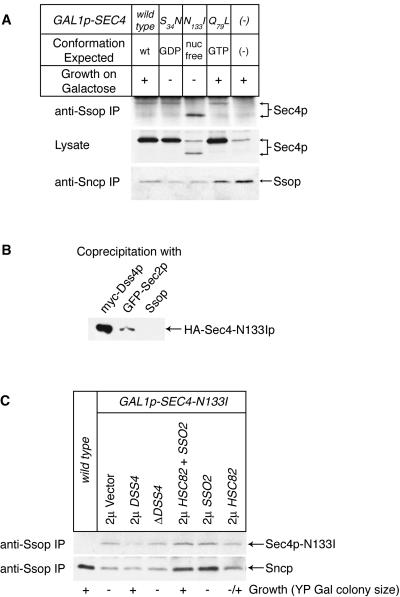
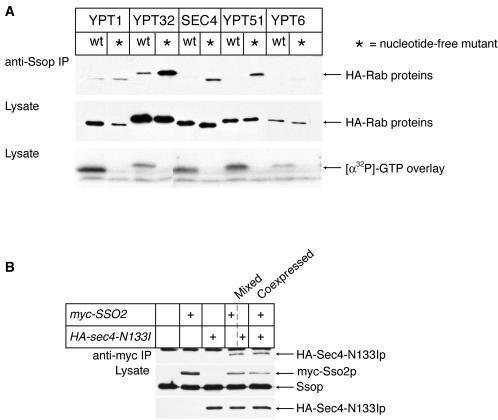
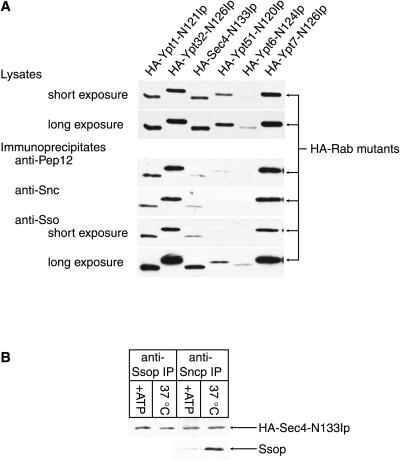
Fusion of post-Golgi secretory vesicles with the plasma membrane in yeast requires the function of a Rab protein, Sec4p, and a set of v- and t-SNAREs, the Snc, Sso, and Sec9 proteins. We have tested the hypothesis that a selective interaction between Sec4p and the exocytic SNAREs is responsible for ensuring that secretory vesicles fuse with the plasma membrane but not with intracellular organelles. Assembly of Sncp and Ssop into a SNARE complex is defective in a sec4-8 mutant strain. However, Snc2p binds in vivo to many other syntaxin-like t-SNAREs, and binding of Sncp to the endosomal/Golgi t-SNARE Tlg2p is also reduced in sec4-8 cells. In addition, binding of Sncp to Ssop is reduced by mutations in two other Rab genes and four non-Rab genes that block the secretory pathway before the formation of secretory vesicles. In an alternate approach to look for selective Rab-SNARE interactions, we report that the nucleotide-free form of Sec4p coimmunoprecipitates with Ssop. However, Rab-SNARE binding is nonselective, because the nucleotide-free forms of six Rab proteins bind with similar low efficiency to three SNARE proteins, Ssop, Pep12p, and Sncp. We conclude that Rabs and SNAREs do not cooperate to specify the target membrane.
Figures
Similar articles
-
Ordering the final events in yeast exocytosis.J Cell Biol. 2000 Oct 16;151(2):439-52. doi: 10.1083/jcb.151.2.439. J Cell Biol. 2000. PMID: 11038189 Free PMC article.
-
Yeast Golgi SNARE interactions are promiscuous.J Cell Sci. 2000 Jan;113 ( Pt 1):145-52. doi: 10.1242/jcs.113.1.145. J Cell Sci. 2000. PMID: 10591633
-
Sec1p binds to SNARE complexes and concentrates at sites of secretion.J Cell Biol. 1999 Jul 26;146(2):333-44. doi: 10.1083/jcb.146.2.333. J Cell Biol. 1999. PMID: 10427089 Free PMC article.
-
How do Rab proteins function in membrane traffic?Int J Biochem Cell Biol. 2000 Mar;32(3):303-7. doi: 10.1016/s1357-2725(99)00112-0. Int J Biochem Cell Biol. 2000. PMID: 10716627 Review.
-
Multisubunit tethering complexes and their role in membrane fusion.Curr Biol. 2010 Nov 9;20(21):R943-52. doi: 10.1016/j.cub.2010.09.015. Curr Biol. 2010. PMID: 21056839 Review.
Cited by
-
Synthetic interactions of the post-Golgi sec mutations of Saccharomyces cerevisiae.Genetics. 2000 Nov;156(3):943-51. doi: 10.1093/genetics/156.3.943. Genetics. 2000. PMID: 11063675 Free PMC article.
-
Pag1p, a novel protein associated with protein kinase Cbk1p, is required for cell morphogenesis and proliferation in Saccharomyces cerevisiae.Mol Biol Cell. 2002 Feb;13(2):503-14. doi: 10.1091/mbc.01-07-0365. Mol Biol Cell. 2002. PMID: 11854408 Free PMC article.
-
Membrane Fusion and SNAREs: Interaction with Ras Proteins.Int J Mol Sci. 2022 Jul 22;23(15):8067. doi: 10.3390/ijms23158067. Int J Mol Sci. 2022. PMID: 35897641 Free PMC article. Review.
-
Ordering the final events in yeast exocytosis.J Cell Biol. 2000 Oct 16;151(2):439-52. doi: 10.1083/jcb.151.2.439. J Cell Biol. 2000. PMID: 11038189 Free PMC article.
-
Distinct trafficking routes of polarized and non-polarized membrane cargoes in Aspergillus nidulans.Elife. 2024 Oct 21;13:e103355. doi: 10.7554/eLife.103355. Elife. 2024. PMID: 39431919 Free PMC article.
References
-
- Abeliovich H, Grote E, Novick P, Ferro-Novick S. Tlg2p, a yeast syntaxin homolog that resides on the Golgi and endocytic structures. J Biol Chem. 1998;273:11719–11727. - PubMed
-
- Botstein D, Segev N, Stearns T, Hoyt MA, Holden J, Kahn RA. Diverse biological functions of small GTP-binding proteins in yeast. Cold Spring Harb Symp Quant Biol. 1988;53:629–636. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
