

Discovery of a novel coronavirus, China Rattus coronavirus HKU24, from Norway rats supports the murine origin of Betacoronavirus 1 and has implications for the ancestor of Betacoronavirus lineage A
- PMID: 25552712
- PMCID: PMC4337523
- DOI: 10.1128/JVI.02420-14
Discovery of a novel coronavirus, China Rattus coronavirus HKU24, from Norway rats supports the murine origin of Betacoronavirus 1 and has implications for the ancestor of Betacoronavirus lineage A
Abstract
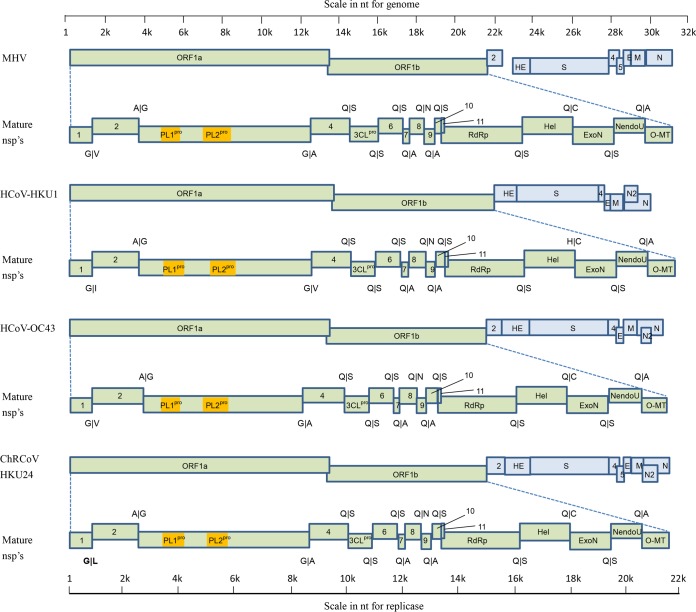
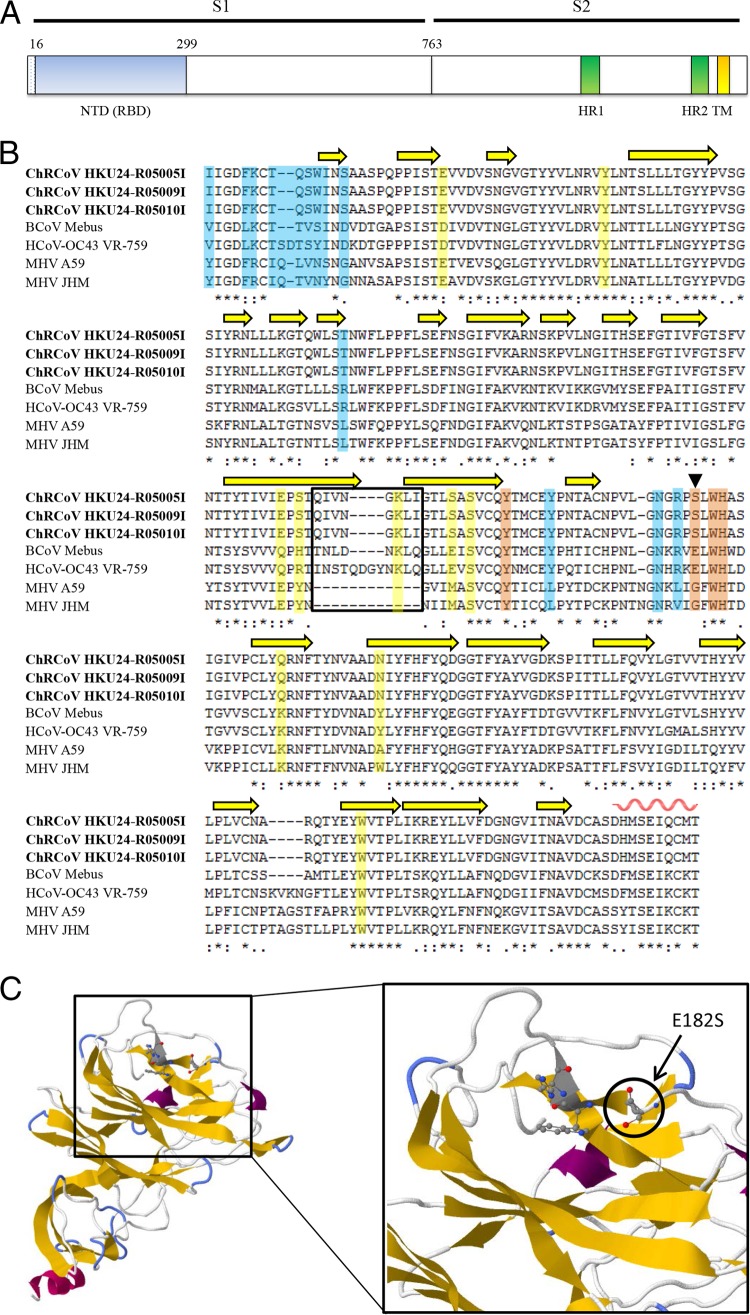
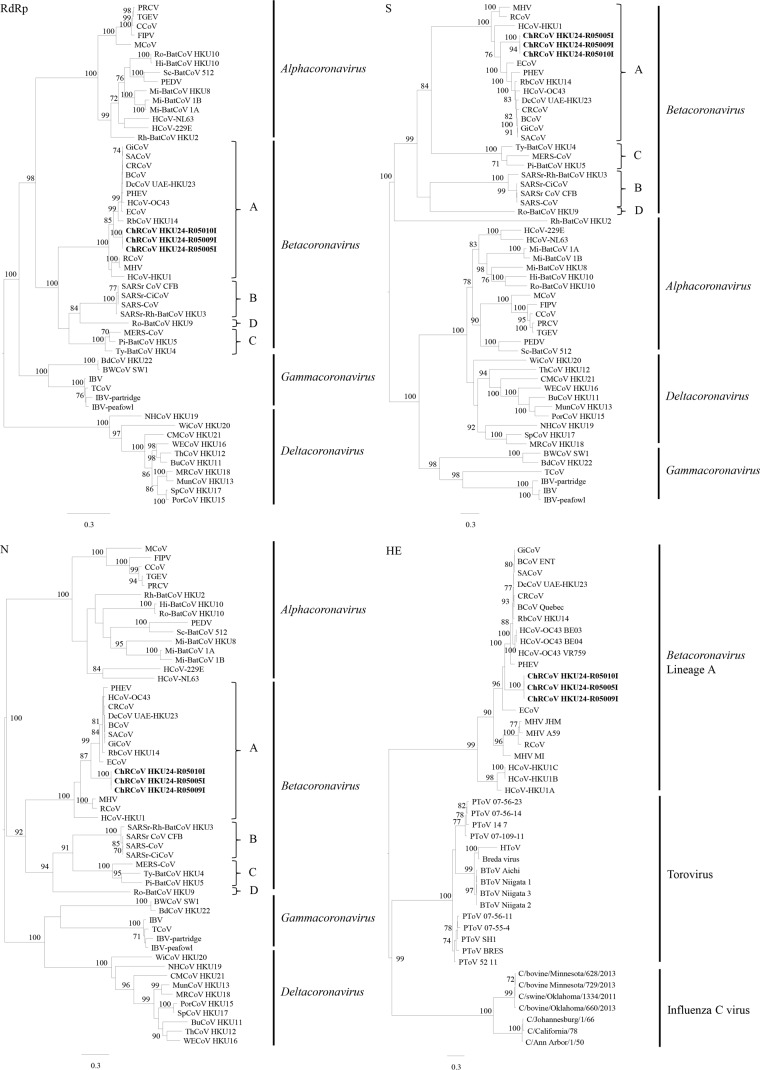
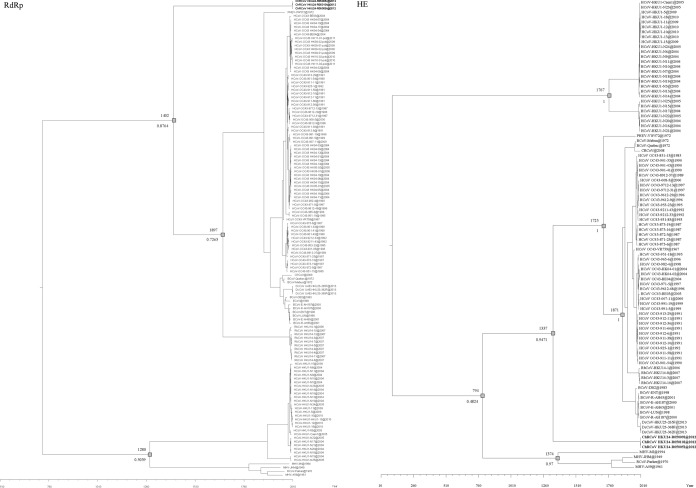
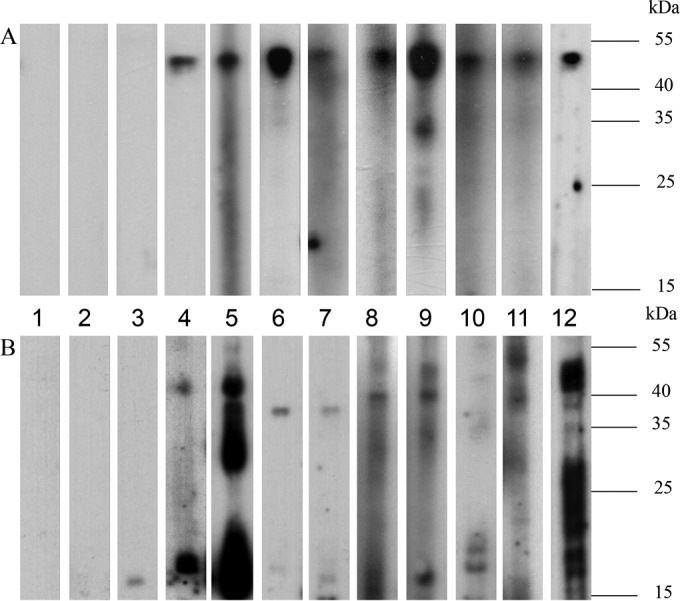
We discovered a novel Betacoronavirus lineage A coronavirus, China Rattus coronavirus (ChRCoV) HKU24, from Norway rats in China. ChRCoV HKU24 occupied a deep branch at the root of members of Betacoronavirus 1, being distinct from murine coronavirus and human coronavirus HKU1. Its unique putative cleavage sites between nonstructural proteins 1 and 2 and in the spike (S) protein and low sequence identities to other lineage A betacoronaviruses (βCoVs) in conserved replicase domains support ChRCoV HKU24 as a separate species. ChRCoV HKU24 possessed genome features that resemble those of both Betacoronavirus 1 and murine coronavirus, being closer to Betacoronavirus 1 in most predicted proteins but closer to murine coronavirus by G+C content, the presence of a single nonstructural protein (NS4), and an absent transcription regulatory sequence for the envelope (E) protein. Its N-terminal domain (NTD) demonstrated higher sequence identity to the bovine coronavirus (BCoV) NTD than to the mouse hepatitis virus (MHV) NTD, with 3 of 4 critical sugar-binding residues in BCoV and 2 of 14 contact residues at the MHV NTD/murine CEACAM1a interface being conserved. Molecular clock analysis dated the time of the most recent common ancestor of ChRCoV HKU24, Betacoronavirus 1, and rabbit coronavirus HKU14 to about the year 1400. Cross-reactivities between other lineage A and B βCoVs and ChRCoV HKU24 nucleocapsid but not spike polypeptide were demonstrated. Using the spike polypeptide-based Western blot assay, we showed that only Norway rats and two oriental house rats from Guangzhou, China, were infected by ChRCoV HKU24. Other rats, including Norway rats from Hong Kong, possessed antibodies only against N protein and not against the spike polypeptide, suggesting infection by βCoVs different from ChRCoV HKU24. ChRCoV HKU24 may represent the murine origin of Betacoronavirus 1, and rodents are likely an important reservoir for ancestors of lineage A βCoVs.
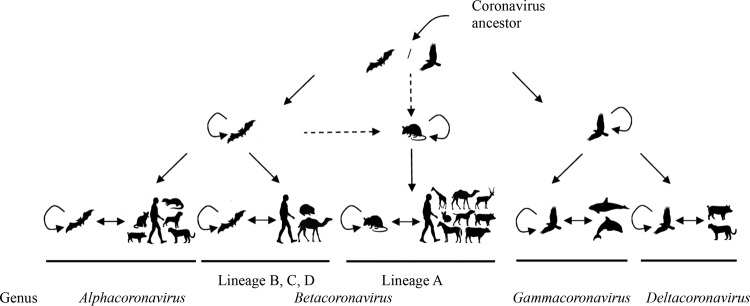
Importance: While bats and birds are hosts for ancestors of most coronaviruses (CoVs), lineage A βCoVs have never been found in these animals and the origin of Betacoronavirus lineage A remains obscure. We discovered a novel lineage A βCoV, China Rattus coronavirus HKU24 (ChRCoV HKU24), from Norway rats in China with a high seroprevalence. The unique genome features and phylogenetic analysis supported the suggestion that ChRCoV HKU24 represents a novel CoV species, occupying a deep branch at the root of members of Betacoronavirus 1 and being distinct from murine coronavirus. Nevertheless, ChRCoV HKU24 possessed genome characteristics that resemble those of both Betacoronavirus 1 and murine coronavirus. Our data suggest that ChRCoV HKU24 represents the murine origin of Betacoronavirus 1, with interspecies transmission from rodents to other mammals having occurred centuries ago, before the emergence of human coronavirus (HCoV) OC43 in the late 1800s. Rodents are likely an important reservoir for ancestors of lineage A βCoVs.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Isolation and characterization of a novel Betacoronavirus subgroup A coronavirus, rabbit coronavirus HKU14, from domestic rabbits.J Virol. 2012 May;86(10):5481-96. doi: 10.1128/JVI.06927-11. Epub 2012 Mar 7. J Virol. 2012. PMID: 22398294 Free PMC article.
-
Identification of the Receptor-Binding Domain of the Spike Glycoprotein of Human Betacoronavirus HKU1.J Virol. 2015 Sep;89(17):8816-27. doi: 10.1128/JVI.03737-14. Epub 2015 Jun 17. J Virol. 2015. PMID: 26085157 Free PMC article.
-
Genetic characterization of Betacoronavirus lineage C viruses in bats reveals marked sequence divergence in the spike protein of pipistrellus bat coronavirus HKU5 in Japanese pipistrelle: implications for the origin of the novel Middle East respiratory syndrome coronavirus.J Virol. 2013 Aug;87(15):8638-50. doi: 10.1128/JVI.01055-13. Epub 2013 May 29. J Virol. 2013. PMID: 23720729 Free PMC article.
-
Bat origin of human coronaviruses.Virol J. 2015 Dec 22;12:221. doi: 10.1186/s12985-015-0422-1. Virol J. 2015. PMID: 26689940 Free PMC article. Review.
-
Hosts and Sources of Endemic Human Coronaviruses.Adv Virus Res. 2018;100:163-188. doi: 10.1016/bs.aivir.2018.01.001. Epub 2018 Feb 16. Adv Virus Res. 2018. PMID: 29551135 Free PMC article. Review.
Cited by
-
Structural basis for mouse receptor recognition by bat SARS2-like coronaviruses.Proc Natl Acad Sci U S A. 2024 Aug 6;121(32):e2322600121. doi: 10.1073/pnas.2322600121. Epub 2024 Jul 31. Proc Natl Acad Sci U S A. 2024. PMID: 39083418 Free PMC article.
-
Molecular determinants of cross-species transmission in emerging viral infections.Microbiol Mol Biol Rev. 2024 Sep 26;88(3):e0000123. doi: 10.1128/mmbr.00001-23. Epub 2024 Jun 24. Microbiol Mol Biol Rev. 2024. PMID: 38912755 Review.
-
Binding affinity between coronavirus spike protein and human ACE2 receptor.Comput Struct Biotechnol J. 2024 Jan 17;23:759-770. doi: 10.1016/j.csbj.2024.01.009. eCollection 2024 Dec. Comput Struct Biotechnol J. 2024. PMID: 38304547 Free PMC article. Review.
-
Human coronavirus HKU1 recognition of the TMPRSS2 host receptor.bioRxiv [Preprint]. 2024 Jan 9:2024.01.09.574565. doi: 10.1101/2024.01.09.574565. bioRxiv. 2024. Update in: Cell. 2024 Aug 8;187(16):4231-4245.e13. doi: 10.1016/j.cell.2024.06.006. PMID: 38260518 Free PMC article. Updated. Preprint.
-
RNA virus diversity in rodents.Arch Microbiol. 2023 Dec 1;206(1):9. doi: 10.1007/s00203-023-03732-4. Arch Microbiol. 2023. PMID: 38038743 Review.
References
-
- de Groot RJ, Baker SC, Baric R, Enjuanes L, Gorbalenya A, Holmes KV, Perlman S, Poon L, Rottier PJ, Talbot PJ, Woo PC, Ziebuhr J. 2011. Coronaviridae, p 806–828 InKing AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (ed), Virus taxonomy: classification and nomenclature of viruses. Ninth report of the International Committee on Taxonomy of Viruses. Academic Press, London, United Kingdom.
-
- Woo PC, Lau SK, Lam CS, Lau CC, Tsang AK, Lau JH, Bai R, Teng JL, Tsang CC, Wang M, Zheng BJ, Chan KH, Yuen KY. 2012. Discovery of seven novel mammalian and avian coronaviruses in Deltacoronavirus supports bat coronaviruses as the gene source of Alphacoronavirus and Betacoronavirus and avian coronaviruses as the gene source of Gammacoronavirus and Deltacoronavirus. J Virol 86:3995–4008. doi:10.1128/JVI.06540-11. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
