

Abstract
Amyotrophic lateral sclerosis (ALS), the most prevalent motor neuron disease characterized by its complex genetic structure, lacks a single diagnostic test capable of providing a conclusive diagnosis. In order to demonstrate the potential for genetic diagnosis and shed light on the pathogenic role of miRNAs in ALS, we developed an ALS diagnostic rule by training the model using 80% of a miRNA profiling dataset consisting of 253 ALS samples and 103 control samples. Subsequently, we validated the diagnostic rule using the remaining 20% of unseen samples. The diagnostic rule we developed includes miR-205-5p, miR-206, miR-376a-5p, miR-412-5p, miR-3927-3p, miR-4701-3p, miR-6763-5p, and miR-6801-3p. Remarkably, the rule achieved an 82% true positive rate and a 73% true negative rate when predicting the unseen samples. Furthermore, the identified miRNAs target 21 genes in the PI3K-Akt pathway and 27 genes in the ALS pathway, including notable genes such as BCL2, NEFH, and OPTN. We propose that miRNA profiling may serve as a complementary diagnostic tool to supplement the clinical presentation and aid in the early recognition of ALS.
Subject terms: Computational biology and bioinformatics, Amyotrophic lateral sclerosis
Introduction
ALS is a neurodegenerative disease that affects the central nervous system, characterized by motor neuron deficit and short life expectancy, but ALS can be challenging to diagnose, particularly in its early stages. Due to its rarity, physicians often consider more common illnesses before considering ALS, which can delay its diagnosis1. To improve early recognition, ALS diagnosis criteria have been proposed2 and continuously revised3. However, even the recently proposed Gold Coast criteria4 is still primarily based on clinical presentation, despite the fact that the genetic structure and biomarkers are gradually revealed by recent studies5. Additionally, newly developed predictive models, scales, and scoring systems can help patients and their physicians better understand the disease course. Although mechanism-based and potentially disease-modifying therapies are currently under clinical trials5, developing new diagnostic criteria and identifying genetic risk factors could also speed up the diagnostic process, and understanding the multisystem nature of ALS, including cognitive dysfunction and behavioral changes, is crucial for providing proper caregiving support and making end-of-life decisions.
ALS is currently divided into familial and sporadic categories. Familial ALS makes up 10–15% of cases and is inherited from family members with ALS or related syndromes like frontotemporal dementia6. About 70% of familial cases have mutations in known ALS genes. On the other hand, sporadic ALS makes up approximately 85% of cases and develops in patients without any family history of ALS. However, around 15% of sporadic ALS cases have private pathogenic mutations in known ALS genes, meaning they do not have a family history of ALS6. The cause of the remaining 85% of sporadic cases is unknown. Over 40 genes linked to ALS have been discovered, each varying in penetrance, frequency, and mode of inheritance. Among these genes, C9orf72, TARDBP, SOD1, and FUS are the most prevalent and have the highest penetrance7. Either toxic gain-of-function or loss-of-function mutations in these known genes are associated with ALS pathological processes. These mutations lead to protein aggregates forming, a key pathological feature in both sporadic and familial ALS cases8. The underlying pathophysiological processes can be broadly categorized into four main types: impaired RNA metabolism, altered proteostasis or autophagy, cytoskeletal or trafficking defects, and mitochondrial dysfunction9. For example, RNA metabolism is often affected by ALS-associated genes such as C9orf72, TARDBP, and FUS. Inclusions of TDP-43 and FUS can impair the normal function of DNA and RNA binding proteins, leading to significant changes in transcription and RNA processing. ALS-associated genes such as TDP-43 can also cause dysregulation of proteostasis and autophagy by preventing damaged protein clearance. Additionally, cytoskeletal and tubulin defects induced by mutant ALS genes like TUBA4A and PFN1 can interfere with axonal trafficking. Finally, SOD1 is a key gene linked to ALS and has been shown to cause mitochondrial dysfunction and increase oxidative stress, which is central to the development of the disease. Despite significant advances, our understanding of the complete molecular basis for the pathophysiology remains incomplete5.
MiRNAs are small non-coding molecules that post-transcriptionally regulate various biological processes, including neuronal function, development, and progression of diseases10. The expression level of miRNAs, including miR-1, miR-9, miR-133a, miR-133a/b, miR-142-3p, miR-149, miR-206, miR-223-3p, miR-326, miR-338-3p, miR-374b-5p, miR-424, and miR-451, in the serum, muscle, tissue, and cerebrospinal fluid of individuals with ALS differs from that of healthy controls11–17. These changes in miRNA expression may target the peripheral nerves, CNS, or skeletal muscles, potentially contributing to the etiology of ALS and playing a crucial role in its progression18. Therefore, Emde, et al., have proposed that different subtypes of ALS may share a common feature: the global dysregulation of miRNAs19. Thus, detecting changes in miRNA expression profile may serve as a valuable diagnostic biomarker to identify disease onset and progression. Additionally, identifying dysregulated miRNAs could offer a promising avenue for developing therapeutic approaches to treat ALS. However, due to ALS's polygenic nature and complexity, no single miRNA has been found universally dysregulated among previous studies. This fact makes it challenging to use miRNA as a diagnostic tool for ALS.
Here, we demonstrate the idea that ALS diagnosis could be assisted with the combination of the expression level of several circulating miRNAs. We established the judgement rule by machine learning using publically available dataset contributed by Magen and colleagues20, and showed that this rule model predicts 82% true positive (ALS) and 73% true negative (healthy control) in the blind dataset, and identified several novel miRNAs that target ALS genes.
Methods
Data source and batch correction
The raw counts of miRNA were downloaded from the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO) database21 of the accession number GSE168714, contributed by Magen and colleagues20. The dataset was accessed at https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE168714 from March 1st to 3rd, 2023. The dataset comprises the annotated counts by small RNA-seq of the RNA extracted from the plasma of 253 ALS and 103 control20. The batch number of data collection and ALS/control state were also available with the raw count. To account for the differences in our study design compared to the original work of Magen and colleagues, we performed our own batch correction using the following method. Firstly, we temporarily excluded highly present miRNAs whose raw count represented more than 2% of the total counts (13 miRNAs excluded, as detailed in Supplementary File 1) during the calculation of batch correction coefficient. Secondly, we calculated the sum of miRNA counts for each sample. Thirdly, we determined the average sum of miRNA counts for each batch. We then defined a batch coefficient as the quotient of the maximum average sum of all batches divided by the average sum of the batch. Finally, we calculated the corrected count, including the temporarily excluded highly present miRNAs, by multiplying the raw counts by the batch coefficient.
Machine learning
The strategy to identify the key miRNAs was adopted from our previous studies22,23. Briefly, the transposition of the corrected miRNA counts served as the input file to the machine learning program RapidMiner24. For clarity, bold italicized text is used to denote the terminology in RapidMiner. In the RapidMiner process setting, patient ID was used as ID; patient ALS state was used as Label, and the corrected miRNA counts were used as Attributes to perform training. Rule Induction was adopted as the algorithm, and the overall training and validation program was shown in Supplementary File 2, with the following parameters. The Split Data operator separated the sample library into 80/20 sets by shuffled sampling for model building and independent validation, respectively. Rule Induction was performed with the criterion of information gain, sample ratio of 0.9, pureness of 0.9, and minimal prune benefit of 0.25, while ten times of Cross Validation was used to improve the model. Finally, the Apply model operator utilized the 20% blind set to validate the generated Rule Model, and the Performance operator demonstrated the performance of the model.
mRNA targets of miRNAs
DIANA-TarBase v7 was used to find the experimentally validated mRNA targets of miRNAs25. Data were accessed at http://diana.imis.athena-innovation.gr/DianaTools/index.php?r=tarbase/index from April 13th to 20th, 2023.
Interaction network and enrichment
String-db version 11.5 was used to generate the interaction network and pathway enrichment26. Data were accessed at https://string-db.org/ from April 20th to 23rd, 2023. Interaction network was generated with highest confidence (0.9) and disconnected nodes in the network were hidden.
Intersection analysis
Venny 2.0 was used to generate the Venn diagram27. Data was accessed at https://bioinfogp.cnb.csic.es/tools/venny/index2.0.2.html from April 20th to 23rd, 2023.
Statistics
Student’s T-test was used to estimate the significance of the difference between two groups.
Results
Machine learning has been proven to be a powerful tool to identify genetic biomarkers in neurodegenerative disorders with complex and heterogeneous genetic factors, such as Alzheimer’s disease22 and Huntington’s disease23. Since ALS is also known for its complexity and heterogeneity genetic architecture, we applied machine learning on an ALS-control miRNA-seq dataset20 to establish a judgment rule compromising miRNA profiles to identify ALS biomarkers following the strategy illustrated in Fig. 1. After batch correction and transposition, batch-corrected counts of miRNAs of 253 ALS and 103 control was generated (Supplementary File 3), and served as the input file for machine learning, where 80% samples served as the training set, and 20% as the unseen testing set. The generated model is shown in Fig. 2A, compromising miR-205-5p, miR-206, miR-376a-5p, miR-412-5p, miR-3927-3p, miR-4701-3p, miR-6763-5p, and miR-6801-3p, and its ROC curve in the training stage is shown in Fig. 2B. This rule model predicts 82% true positive and 73% true negative in the unseen dataset. The expression fold change of the identified miRNAs is shown in Fig. 3, with miR-412-5p, miR-3927-3p, miR-4701-3p, and miR-6801-3p significantly down-regulated in ALS, but not the other four miRNAs. This reflects the idea that a major advance of machine learning over traditional comparative methods in identifying biomarkers from expression profiles22.
Figure 1.
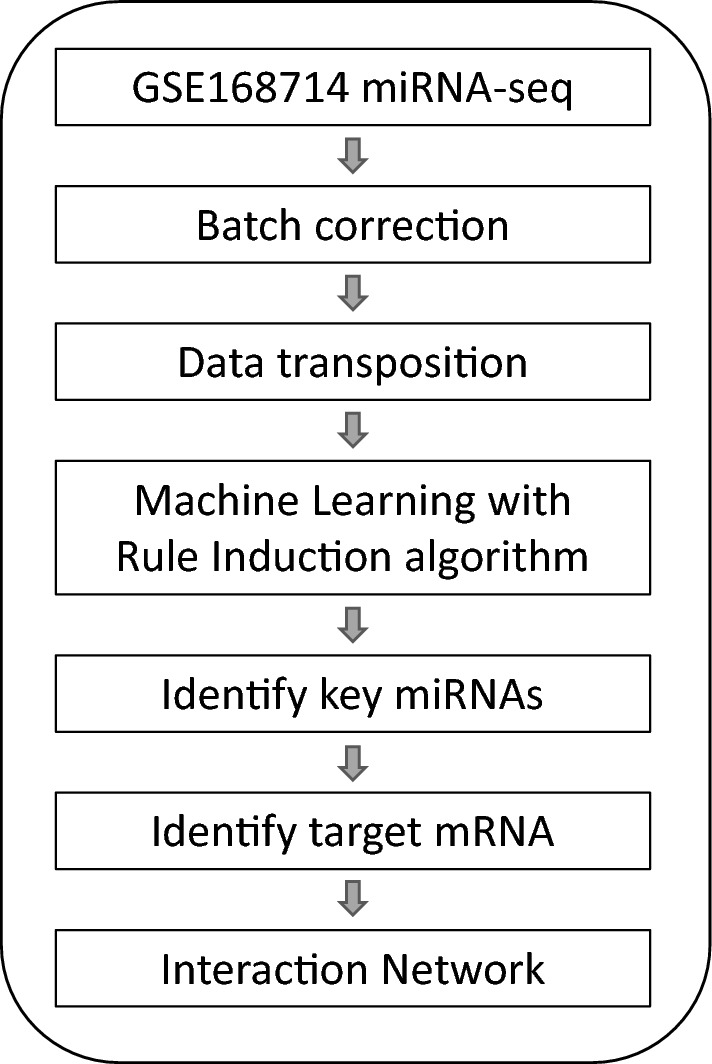
The workflow of this study.
Figure 2.

The predictive model generated by machine learning and its performance. (A) The rule model judging where a sample is ALS or control, where “1” denotes ALS and “0” denotes control. The value in the conditional expression is the batch-corrected count of the miRNA. (B) The ROC curve of the rule model, with AUC of 0.831.
Figure 3.

The miRNA expression fold change of the miRNAs identified by the rule model. Error bars stand for the standard error of the mean (SEM); while *, **, and *** stand for p-value < 0.05, 0.01, and 0.001 of Student’s T-test, respectively. The upper right panel shows the number of experimentally validated mRNA targets of miRNA, with the full list provided in the corresponding supplementary files.
To understand the role of miRNA-mediated biological processes in ALS pathology, we inquired the mRNA targets of miR-205-5p, miR-206, miR-376a-5p, miR-412-5p, miR-3927-3p, miR-4701-3p, miR-6763-5p, and miR-6801-3p in TarBase25, a curated database of miRNA-target interactions experimental evidence. Four miRNAs, i.e., miR-3927-3p, miR-4701-3p, miR-6763-5p, and miR-6801-3p, have no experiment-validated targets, while miR-205-5p, miR-206, miR-376a-5p, miR-412 have 296, 100, 43, and 200 targets, respectively (Supplementary File 4–7). Next, we showed that miRNA target genes enrich ALS and PI3K-Akt signaling pathways (Fig. 4, Supplementary File 8, 9), forming an interaction network with miR-205-5p targets occupying central hubs. Interestingly, The Venn diagram shows that the miRNA-target sets are nearly mutually exclusive in both ALS (Fig. 5A) and PI3K-Akt signaling pathway (Fig. 5B).
Figure 4.

The protein interaction network of the mRNA targets of the identified miRNAs. Red balls indicate proteins in the Amyotrophic lateral sclerosis of KEGG, while blue balls indicate proteins in the PI3K-Akt signaling pathway of KEGG.
Figure 5.

The Venn diagram showing that the miRNA-target sets are nearly mutually exclusive. (A) The ALS genes. (B) The PI3K-Akt genes.
Discussion
In this study, we proposed a diagnostic rule of ALS by the expression of miRNAs, including miR-205-5p, miR-206, miR-376a-5p, miR-412-5p, miR-3927-3p, miR-4701-3p, miR-6763-5p, and miR-6801-3p, where miR-206 has been reported in several previous ALS studies, while other miRNAs are novel in the ALS research field. Furthermore, these miRNAs mutually exclusively target genes in the ALS or PI3K-Akt pathways, which supports the idea that the combination of miRNAs, but not any single miRNA, may serve as a tool to facilitate ALS diagnosis. In the context of ALS’s heterogeneous genetics, we discuss the identified miRNAs’ and their target genes’ relevance to ALS below.
Heterogeneity of ALS genetics
Recent advancements have greatly enhanced our comprehension of the genetic origins of familial ALS. Approximately 40–55% of familial ALS cases can be accounted for by variations in well-known ALS-linked genes7. While more than 50 potential causative or disease-modifying genes have been identified, pathogenic variants in SOD1, C9ORF72, FUS, and TARDBP are most frequent, whereas variants in other genes are relatively uncommon28. However, in cases of sporadic ALS, diagnostic progress has only elucidated a fraction of the cases, with the etiology remaining unexplained in over 90% of patients29. It is widely accepted that genetic risk factors play a significant role in sporadic ALS, with heritability estimated at approximately 60% based on twin studies30. However, despite extensive genetic association studies, identifying heritable genetic risk factors in sporadic ALS remains elusive.
Despite decades of research, ALS's underlying causative pathogenic mechanisms remain uncertain, particularly in sporadic cases. The development and progression of the disease are likely influenced by multiple factors rather than a single initiating event31. Moreover, genetic and phenotypic variations among patients pose challenges in comprehending and drawing conclusions about the general pathogenic mechanisms of ALS. Given the extensive number of genes and cellular processes implicated in ALS, numerous disease mechanisms have been proposed, including disruptions in RNA metabolism32, compromised protein homeostasis33, defects in nucleocytoplasmic transport34, impaired DNA repair35, mitochondrial dysfunction36, oxidative stress37, disturbances in axonal transport38, and oligodendrocyte dysfunction39. Further clarification is required to determine the timing and extent to which each of these mechanisms contributes to the pathogenesis of ALS.
As an attempt to see whether ALS clinical phenotypes could be differentiated by miRNAs, we constructed new cohorts of ALS patients from the same GSE168714 dataset by their clinical phenotypes: bulbar-onset or non-bulbar-onset, with 83 or 170 patients, respectively (Supplementary File 10), and these new cohorts of patients were subjected to de novo analysis in machine learning. However, the performance of the newly-established model was poor, with a recall rate of 75.9% and 24.6% for the cohorts, respectively. Thus, machine learning could not differentiate the cohorts of ALS, at least under the present condition. We further analyzed whether the identified miRNAs express differentially in the cohorts of ALS (Supplementary File 11). We found that miR-206 and miR-205 express differentially in the cohorts of ALS, but not significantly. We suspect that the sample size is the bottleneck to uncover the nature of the ALS cohorts. We also summarized other publicly-available miRNA datasets (Supplementary File 12). However, their sample size is insufficient for an independent machine learning study, and none of the datasets meets the requirement for validation of the current model that all eight miRNAs used in the machine learning model must be available.
miR-206
miR-206 participates in various stages of muscle differentiation, encompassing alternative splicing, DNA synthesis, and cell apoptosis40. During development, miR-206 hinders the activity of Pax7 and Pax3, effectively restricting the proliferative potential of satellite cells while promoting their differentiation into myogenic progenitor cells. Conversely, reducing miR-206 leads to the overexpression of Pax7 and Pax3, which consequently inhibits the differentiation of myoblasts. As Pax7 and Pax3 are known pro-survival factors, the down-regulation of miR-206 can induce apoptosis41. Thus, miR-206 plays a protective role and facilitates the regeneration of neuromuscular junctions following acute nerve injury, particularly in the context of ALS42. Knock-out of miR-206 delays and mutilates muscle reinnervation in ALS mouse models of SOD mutant43. Recent findings indicate elevated levels of miR-206 in the plasma of ALS patients and could indicate of disease progression44,45.
miR-205
miR-205 exhibits significant expression levels in various human epithelial tissues, including the breast, prostate, skin, eye, and thymus. Its primary role in these tissues is crucial to tissue morphogenesis and homeostasis. Specifically, it upholds the epithelial phenotype by directly targeting two transcription factors: zinc finger E-box-binding homeobox 1 (ZEB1) and ZEB2, which repress E-cadherin and other genes associated with polarity46. During the early stages of embryonic development, miR-205 is expressed in trophoblasts, where it regulates placental development by suppressing the Mediator of RNA polymerase II transcription subunit 1 (MED1)47. Moreover, in embryonic development, miR-205 governs the differentiation of extraembryonic endoderm and spermatogenesis by influencing cell migration and adhesion genes48. In the mammary gland, miR-205 displays high expression levels in the basal stem cells. Overexpression of miR-205 has been shown to induce the expansion of the progenitor cell population while reducing cell size and promoting cellular proliferation. These effects are achieved by repressing PTEN49. In this context, miR-205 regulates the production of the basement membrane protein complex laminin-332 and its receptor integrin-β4, thereby ensuring proper tissue polarity and morphogenesis50. In the skin epidermis and stratified epithelia of the esophagus and tongue, miR-205 has been found to play a significant role in expanding the stem cell population through its regulation of PI3K signaling51. Additionally, by influencing the same signaling pathways, miR-205 enhances the migration of human epidermal and corneal epithelial keratinocytes, thereby contributing to wound healing and corneal development52.
miR-376a and miR-412
The physiological roles of miR-376a and miR-412 are not fully understood yet, despite some studies reported its participation in cancer and neurological disorders. For example, increased levels of miR-376a have been observed in the T cells of patients with multiple sclerosis (MS)53, and miR-412 may inhibit clear cell renal cell carcinoma progression54. Conversely, in the late-onset form of Alzheimer's disease (LOAD), miR-376a has been identified as down-regulated in the brain55. Meanwhile, the expression change of miR-412 has been mentioned in the brain of alcohol use disorder56, and also in Alzheimer's disease57.
BCL2
BCL2 is targeted by miR-20558 and controls caspase activation and the initiation of programmed cell death59 and thus regulates neuronal development and neurodegeneration60. Several lines of evidence show that BCL2 probably involves in ALS pathological progression. Epidemiologic studies found altered expression of BCL2 in ALS spinal cord motor neurons61 and post‐central gyrus62. In vivo studies showed that overexpression of BCL2 prolongs the survival of the ALS mouse model63 and improves neuromuscular function64. In vitro studies revealed that ALS-associated mutant SOD1 aggregates BCL265 and advocates BCL2 conformational changes66.
NEFH
Neurofilament heavy polypeptide (NEFH) is one of the three intermediate filament proteins forming neurofilaments67. NEFH is targeted by miR-20568 and could be phosphorylated by GSK3β69 and regulate the Akt-β-catenin pathway70. Moreover, epidemiological study showed that NEFH mutation71,72 and expression73 is associated with ALS. Besides, NEFH mutation or expression is associated with other disorders of central or peripheral neural system, including schizophrenia74, alcoholics75, and Charcot-Marie-Tooth neuropathy76.
OPTN
OPTN, also known as optineurin, is a highly conserved protein in various species77. OPTN is target by miR-20568 as well. It plays diverse roles in vesicular trafficking, NFKB/NF-κB signaling, and autophagy. Specifically, OPTN has been identified as an autophagy receptor that facilitates the connection between ubiquitinated autophagy substrates and MAP1LC3/LC3-positive phagophore membranes78. Furthermore, mounting evidence suggests that OPTN acts as an inducer of autophagy, initiating the autophagic process79,80. Moreover, studies indicate that OPTN's involvement in autophagic initiation can commence as early as the formation of autophagosomal membranes81,82. These groundbreaking findings underscore the multifunctional role of OPTN as a potential autophagy receptor throughout the autophagic process, expanding beyond its traditional perception as a receptor operating solely at a single stage of autophagy. OPTN is gathering attention in ALS research, since variants of this gene are associated with ALS83–85. Moreover, OPTN mutant induces neuronal cell death by mediating mitophagy86, autophagy and ER stress87. Interestingly, OPTN mutation might be the common cause of ALS and corticobasal syndrome (CBS)88.
PI3K in ALS
The PI3K-askt signaling pathway governs metabolism, cell survival, motility, transcription, and cell-cycle progression. In recent years, studies have revealed the involvement of the PI3K-Akt signaling pathway in neurodegenerative diseases. For instance, butylphthalide has been shown to activate the PI3K-Akt/GSK-3β signaling pathway in an ischemic cerebral infarction model, reducing nerve function damage and protecting local nerve cells89. Therefore, therapeutic strategies for ALS targeting the PI3K-Akt pathway has been shown to increase anti-apoptotic protein expression levels, reduce pro-apoptotic protein expression levels, and improve cell survival rate and mitochondrial function in ALS90. Moreover, studies by Xiang and colleagues have found that AEG-1 can regulate the PI3K-Akt pathway91, and the absence of AEG-1 in ALS motor neurons inhibits the PI3K-Akt pathway and increases cell apoptosis91. Thus, dysregulated miRNAs may promote ALS pathology by mediating PI3K-Akt signaling pathway.
Conclusion
In sum, we showed that a set of miRNA expressions could serve as a diagnostic tool for ALS, and these miRNAs target ALS and PI3K-Akt pathways in a mutually exclusive way. The key miRNAs include miR-205-5p, miR-206, and miR-376a-5p, while key targets are BCL2, NEFH, and OPTN. We propose that miRNA profiling may facilitate clinical presentation in the early recognition of ALS.
Supplementary Information
Author contributions
W.Y.L. and H.P.L. initiated and supervised this study. J.C. and W.K.H. contributed to the acquisition, analysis, and interpretation of data. B.T.W. contributed to data analysis. All authors discussed and drafted the manuscript.
Funding
This work was supported by grants from National Science and Technology Council of Taiwan (MOST 111-2320-B-039-032, MOST 111-2314-B-039-017-MY3) and grants from China Medical University & Hospital (CMU109-MF-110, CMU111-MF-50, CMU110-MF-92, DMR-112-125).
Data availability
All data in this study are included in the supplementary data. The raw data used for machine learning is shown in Supplementary File 3, and the first two rows (file descriptions) must be removed before use. The raw counts of miRNA are also available from the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO) database of the accession number GSE168714.
Code availability
The machine learning platform RapidMiner Studio is available at https://rapidminer.com/. The process is shown in Supplementary File 2, and the parameters are in the “Methods” section.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Jack Cheng and Wen-Kuang Ho.
Contributor Information
Hsin-Ping Liu, Email: hpliu@mail.cmu.edu.tw.
Wei-Yong Lin, Email: linwy@mail.cmu.edu.tw.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-023-40879-y.
References
- 1.Ryan M, Heverin M, McLaughlin RL, Hardiman O. Lifetime risk and heritability of amyotrophic lateral sclerosis. JAMA Neurol. 2019;76:1367–1374. doi: 10.1001/jamaneurol.2019.2044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Brooks, B. R. El Escorial World Federation of Neurology criteria for the diagnosis of amyotrophic lateral sclerosis. Subcommittee on Motor Neuron Diseases/Amyotrophic Lateral Sclerosis of the World Federation of Neurology Research Group on Neuromuscular Diseases and the El Escorial" Clinical limits of amyotrophic lateral sclerosis" workshop contributors. J. Neurol. Sci.124, 96–107 (1994). [DOI] [PubMed]
- 3.Brooks BR, Miller RG, Swash M, El Munsat TL. Escorial revisited: revised criteria for the diagnosis of amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Other Motor Neuron Disord. 2000;1:293–299. doi: 10.1080/146608200300079536. [DOI] [PubMed] [Google Scholar]
- 4.Shefner JM, et al. A proposal for new diagnostic criteria for ALS. Clin. Neurophysiol. 2020;131:1975–1978. doi: 10.1016/j.clinph.2020.04.005. [DOI] [PubMed] [Google Scholar]
- 5.Feldman EL, et al. Amyotrophic lateral sclerosis. Lancet. 2022;400:1363. doi: 10.1016/S0140-6736(22)01272-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Goutman SA, et al. Emerging insights into the complex genetics and pathophysiology of amyotrophic lateral sclerosis. Lancet Neurol. 2022;21:465. doi: 10.1016/S1474-4422(21)00414-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zou Z-Y, et al. Genetic epidemiology of amyotrophic lateral sclerosis: A systematic review and meta-analysis. J. Neurol. Neurosurg. Psychiatry. 2017;88:540–549. doi: 10.1136/jnnp-2016-315018. [DOI] [PubMed] [Google Scholar]
- 8.Mackenzie IR, Rademakers R, Neumann M. TDP-43 and FUS in amyotrophic lateral sclerosis and frontotemporal dementia. Lancet Neurol. 2010;9:995–1007. doi: 10.1016/S1474-4422(10)70195-2. [DOI] [PubMed] [Google Scholar]
- 9.Nguyen HP, Van Broeckhoven C, van der Zee J. ALS genes in the genomic era and their implications for FTD. Trends Genet. 2018;34:404–423. doi: 10.1016/j.tig.2018.03.001. [DOI] [PubMed] [Google Scholar]
- 10.Cai Y, Yu X, Hu S, Yu J. A brief review on the mechanisms of miRNA regulation. Genomics Proteomics Bioinform. 2009;7:147–154. doi: 10.1016/S1672-0229(08)60044-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Russell AP, et al. Disruption of skeletal muscle mitochondrial network genes and miRNAs in amyotrophic lateral sclerosis. Neurobiol. Dis. 2013;49:107–117. doi: 10.1016/j.nbd.2012.08.015. [DOI] [PubMed] [Google Scholar]
- 12.De Felice, B. et al. miR-338-3p is over-expressed in blood, CFS, serum and spinal cord from sporadic amyotrophic lateral sclerosis patients. Neurogenetics15, 243–253 (2014). [DOI] [PubMed]
- 13.Sumitha R, et al. Differential expression of microRNA-206 in the gastrocnemius and biceps brachii in response to CSF from sporadic amyotrophic lateral sclerosis patients. J. Neurol. Sci. 2014;345:254–256. doi: 10.1016/j.jns.2014.07.005. [DOI] [PubMed] [Google Scholar]
- 14.Hawley ZC, Campos-Melo D, Strong MJ. Novel miR-b2122 regulates several ALS-related RNA-binding proteins. Mol. Brain. 2017;10:1–14. doi: 10.1186/s13041-017-0326-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Pegoraro V, Merico A, Angelini C. Micro-RNAs in ALS muscle: Differences in gender, age at onset and disease duration. J. Neurol. Sci. 2017;380:58–63. doi: 10.1016/j.jns.2017.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Saucier D, et al. Identification of a circulating miRNA signature in extracellular vesicles collected from amyotrophic lateral sclerosis patients. Brain Res. 2019;1708:100–108. doi: 10.1016/j.brainres.2018.12.016. [DOI] [PubMed] [Google Scholar]
- 17.Banack SA, Dunlop RA, Stommel EW, Mehta P, Cox PA. miRNA extracted from extracellular vesicles is a robust biomarker of amyotrophic lateral sclerosis. J. Neurol. Sci. 2022;442:120396. doi: 10.1016/j.jns.2022.120396. [DOI] [PubMed] [Google Scholar]
- 18.Liu H, et al. Systematic review and meta-analysis on microRNAs in amyotrophic lateral sclerosis. Brain Res. Bull. 2023;194:83. doi: 10.1016/j.brainresbull.2023.01.005. [DOI] [PubMed] [Google Scholar]
- 19.Emde A, et al. Dysregulated mi RNA biogenesis downstream of cellular stress and ALS-causing mutations: A new mechanism for ALS. EMBO J. 2015;34:2633–2651. doi: 10.15252/embj.201490493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Magen I, et al. Circulating miR-181 is a prognostic biomarker for amyotrophic lateral sclerosis. Nat. Neurosci. 2021;24:1534–1541. doi: 10.1038/s41593-021-00936-z. [DOI] [PubMed] [Google Scholar]
- 21.Barrett T, et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2012;41:D991–D995. doi: 10.1093/nar/gks1193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cheng J, Liu H-P, Lin W-Y, Tsai F-J. Machine learning compensates fold-change method and highlights oxidative phosphorylation in the brain transcriptome of Alzheimer’s disease. Sci. Rep.-UK. 2021;11:13704. doi: 10.1038/s41598-021-93085-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cheng J, Liu H-P, Lin W-Y, Tsai F-J. Identification of contributing genes of Huntington’s disease by machine learning. BMC Med. Genomics. 2020;13:1–11. doi: 10.1186/s12920-020-00822-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ristoski P, Bizer C, Paulheim H. Mining the web of linked data with rapidminer. J. Web Semantics. 2015;35:142–151. doi: 10.1016/j.websem.2015.06.004. [DOI] [Google Scholar]
- 25.Vlachos IS, et al. DIANA-TarBase v7.0: Indexing more than half a million experimentally supported miRNA: mRNA interactions. Nucleic Acids Res. 2015;43:D153–D159. doi: 10.1093/nar/gku1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Szklarczyk D, et al. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021;49:D605–D612. doi: 10.1093/nar/gkaa1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Oliveros, J. C. Venny. An Interactive Tool for Comparing Lists with Venn's Diagrams. (2007).
- 28.Boylan K. Familial amyotrophic lateral sclerosis. Neurol. Clin. 2015;33:807–830. doi: 10.1016/j.ncl.2015.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Renton AE, Chiò A, Traynor BJ. State of play in amyotrophic lateral sclerosis genetics. Nat. Neurosci. 2014;17:17–23. doi: 10.1038/nn.3584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Al-Chalabi A, et al. An estimate of amyotrophic lateral sclerosis heritability using twin data. J. Neurol. Neurosurg. Psychiatry. 2010;81:1324–1326. doi: 10.1136/jnnp.2010.207464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mejzini R, et al. ALS genetics, mechanisms, and therapeutics: Where are we now? Front. Neurosci. 2019;13:1310. doi: 10.3389/fnins.2019.01310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mandrioli, J., Mediani, L., Alberti, S. & Carra, S. Seminars in Cell & Developmental Biology. 183–192 (Elsevier). [DOI] [PubMed]
- 33.Thompson AG, et al. CSF extracellular vesicle proteomics demonstrates altered protein homeostasis in amyotrophic lateral sclerosis. Clin. Proteomics. 2020;17:1–12. doi: 10.1186/s12014-020-09294-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lin Y-C, et al. Interactions between ALS-linked FUS and nucleoporins are associated with defects in the nucleocytoplasmic transport pathway. Nat. Neurosci. 2021;24:1077–1088. doi: 10.1038/s41593-021-00859-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lopez-Gonzalez R, et al. Partial inhibition of the overactivated Ku80-dependent DNA repair pathway rescues neurodegeneration in C9ORF72-ALS/FTD. Proc. Natl. Acad. Sci. 2019;116:9628–9633. doi: 10.1073/pnas.1901313116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cozzolino M, Carrì MT. Mitochondrial dysfunction in ALS. Prog. Neurobiol. 2012;97:54–66. doi: 10.1016/j.pneurobio.2011.06.003. [DOI] [PubMed] [Google Scholar]
- 37.Barber SC, Shaw PJ. Oxidative stress in ALS: Key role in motor neuron injury and therapeutic target. Free Radical Biol. Med. 2010;48:629–641. doi: 10.1016/j.freeradbiomed.2009.11.018. [DOI] [PubMed] [Google Scholar]
- 38.Bilsland LG, et al. Deficits in axonal transport precede ALS symptoms in vivo. Proc. Natl. Acad. Sci. 2010;107:20523–20528. doi: 10.1073/pnas.1006869107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Boillée S, et al. Onset and progression in inherited ALS determined by motor neurons and microglia. Science. 2006;312:1389–1392. doi: 10.1126/science.1123511. [DOI] [PubMed] [Google Scholar]
- 40.Ma G, et al. MiR-206, a key modulator of skeletal muscle development and disease. Int. J. Biol. Sci. 2015;11:345. doi: 10.7150/ijbs.10921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Chen J-F, et al. microRNA-1 and microRNA-206 regulate skeletal muscle satellite cell proliferation and differentiation by repressing Pax7. J. Cell Biol. 2010;190:867–879. doi: 10.1083/jcb.200911036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Williams AH, et al. MicroRNA-206 delays ALS progression and promotes regeneration of neuromuscular synapses in mice. Science. 2009;326:1549–1554. doi: 10.1126/science.1181046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.De Felice B, et al. A miRNA signature in leukocytes from sporadic amyotrophic lateral sclerosis. Gene. 2012;508:35–40. doi: 10.1016/j.gene.2012.07.058. [DOI] [PubMed] [Google Scholar]
- 44.de Andrade HM, et al. MicroRNAs-424 and 206 are potential prognostic markers in spinal onset amyotrophic lateral sclerosis. J. Neurol. Sci. 2016;368:19–24. doi: 10.1016/j.jns.2016.06.046. [DOI] [PubMed] [Google Scholar]
- 45.Waller R, et al. Serum miRNAs miR-206, 143-3p and 374b-5p as potential biomarkers for amyotrophic lateral sclerosis (ALS) Neurobiol. Aging. 2017;55:123–131. doi: 10.1016/j.neurobiolaging.2017.03.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gregory PA, et al. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell Biol. 2008;10:593–601. doi: 10.1038/ncb1722. [DOI] [PubMed] [Google Scholar]
- 47.Mouillet JF, Chu T, Nelson DM, Mishima T, Sadovsky Y. MiR-205 silences MED1 in hypoxic primary human trophoblasts. FASEB J. 2010;24:2030–2039. doi: 10.1096/fj.09-149724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Li C, Finkelstein D, Sherr CJ. Arf tumor suppressor and miR-205 regulate cell adhesion and formation of extraembryonic endoderm from pluripotent stem cells. Proc. Natl. Acad. Sci. 2013;110:E1112–E1121. doi: 10.1073/pnas.1302184110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Greene SB, Gunaratne PH, Hammond SM, Rosen JM. A putative role for microRNA-205 in mammary epithelial cell progenitors. J. Cell Sci. 2010;123:606–618. doi: 10.1242/jcs.056812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Gandellini P, et al. miR-205 regulates basement membrane deposition in human prostate: Implications for cancer development. Cell Death Differ. 2012;19:1750–1760. doi: 10.1038/cdd.2012.56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wang D, et al. MicroRNA-205 controls neonatal expansion of skin stem cells by modulating the PI (3) K pathway. Nat. Cell Biol. 2013;15:1153–1163. doi: 10.1038/ncb2827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Yu J, et al. MicroRNA-205 promotes keratinocyte migration via the lipid phosphatase SHIP2. FASEB J. 2010;24:3950. doi: 10.1096/fj.10-157404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Nateghi B, Emadi F, Eghbali M, Pezeshki P, Eshaghiyan A. Circulating miR-193b-3p and miR-376a-3p involved in Iranian patients with multiple sclerosis. Int. Biol. Biomed. J. 2019;5:24–28. [Google Scholar]
- 54.Wang Y, et al. Circular RNA circDVL1 inhibits clear cell renal cell carcinoma progression through the miR-412-3p/PCDH7 axis. Int. J. Biol. Sci. 2022;18:1491. doi: 10.7150/ijbs.69351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lau P, Frigerio CS, De Strooper B. Variance in the identification of microRNAs deregulated in Alzheimer's disease and possible role of lincRNAs in the pathology: The need of larger datasets. Ageing Res. Rev. 2014;17:43–53. doi: 10.1016/j.arr.2014.02.006. [DOI] [PubMed] [Google Scholar]
- 56.Lim Y, et al. Exploration of alcohol use disorder-associated brain miRNA–mRNA regulatory networks. Transl. Psychiatry. 2021;11:504. doi: 10.1038/s41398-021-01635-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kumar S, Reddy PH. Are circulating microRNAs peripheral biomarkers for Alzheimer's disease? Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2016;1862:1617–1627. doi: 10.1016/j.bbadis.2016.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Skalsky RL, et al. The viral and cellular microRNA targetome in lymphoblastoid cell lines. PLoS Pathog. 2012;8:e1002484. doi: 10.1371/journal.ppat.1002484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Cory S, Adams JM. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer. 2002;2:647–656. doi: 10.1038/nrc883. [DOI] [PubMed] [Google Scholar]
- 60.Akhtar, R. S., Ness, J. M. & Roth, K. A. Bcl-2 family regulation of neuronal development and neurodegeneration. Biochim. Biophys. Acta (BBA)-Mol. Cell Res.1644, 189–203 (2004). [DOI] [PubMed]
- 61.Mu X, He J, Anderson DW, Springer JE, Trojanowski JQ. Altered expression of bcl-2 and bax mRNA in amyotrophic lateral sclerosis spinal cord motor neurons. Ann. Neurol. 1996;40:379–386. doi: 10.1002/ana.410400307. [DOI] [PubMed] [Google Scholar]
- 62.Troost, D., Aten, J., Morsink, F. & De Jong, J. Apoptosis in amyotrophic lateral sclerosis is not restricted to motor neurons. Bcl‐2 expression is increased in unaffected post‐central gyrus. Neuropathol. Appl. Neurobiol.21, 498–504 (1995). [DOI] [PubMed]
- 63.Kostic V, Jackson-Lewis V, de Bilbao F, Dubois-Dauphin M, Przedborski S. Bcl-2: Prolonging life in a transgenic mouse model of familial amyotrophic lateral sclerosis. Science. 1997;277:559–563. doi: 10.1126/science.277.5325.559. [DOI] [PubMed] [Google Scholar]
- 64.Azzouz M, et al. Increased motoneuron survival and improved neuromuscular function in transgenic ALS mice after intraspinal injection of an adeno-associated virus encoding Bcl-2. Hum. Mol. Genet. 2000;9:803–811. doi: 10.1093/hmg/9.5.803. [DOI] [PubMed] [Google Scholar]
- 65.Pasinelli P, et al. Amyotrophic lateral sclerosis-associated SOD1 mutant proteins bind and aggregate with Bcl-2 in spinal cord mitochondria. Neuron. 2004;43:19–30. doi: 10.1016/j.neuron.2004.06.021. [DOI] [PubMed] [Google Scholar]
- 66.Pedrini S, et al. ALS-linked mutant SOD1 damages mitochondria by promoting conformational changes in Bcl-2. Hum. Mol. Genet. 2010;19:2974–2986. doi: 10.1093/hmg/ddq202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lees J, Shneidman P, Skuntz S, Carden M, Lazzarini R. The structure and organization of the human heavy neurofilament subunit (NF-H) and the gene encoding it. EMBO J. 1988;7:1947–1955. doi: 10.1002/j.1460-2075.1988.tb03032.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Xie, H., Zhao, Y., Caramuta, S., Larsson, C. & Lui, W.-O. miR-205 Expression Promotes Cell Proliferation and Migration of Human Cervical Cancer Cells. (2012). [DOI] [PMC free article] [PubMed]
- 69.Sasaki T, et al. In vivo and in vitro phosphorylation at Ser-493 in the glutamate (E)-segment of neurofilament-H subunit by glycogen synthase kinase 3β. J. Biol. Chem. 2002;277:36032–36039. doi: 10.1074/jbc.M206674200. [DOI] [PubMed] [Google Scholar]
- 70.Kim MS, et al. Neurofilament heavy polypeptide regulates the Akt-β-catenin pathway in human esophageal squamous cell carcinoma. PLoS ONE. 2010;5:e9003. doi: 10.1371/journal.pone.0009003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Lin F, et al. Sequencing of neurofilament genes identified NEFH Ser787Arg as a novel risk variant of sporadic amyotrophic lateral sclerosis in Chinese subjects. BMC Med. Genomics. 2021;14:1–8. doi: 10.1186/s12920-021-01073-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Theunissen F, et al. Intronic NEFH variant is associated with reduced risk for sporadic ALS and later age of disease onset. Sci. Rep.-UK. 2022;12:14739. doi: 10.1038/s41598-022-18942-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Campos-Melo D, Hawley ZC, Strong MJ. Dysregulation of human NEFM and NEFH mRNA stability by ALS-linked miRNAs. Mol. Brain. 2018;11:1–9. doi: 10.1186/s13041-018-0386-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Pinacho R, et al. Altered CSNK1E, FABP4 and NEFH protein levels in the dorsolateral prefrontal cortex in schizophrenia. Schizophr. Res. 2016;177:88–97. doi: 10.1016/j.schres.2016.04.050. [DOI] [PubMed] [Google Scholar]
- 75.Iwamoto K, et al. Decreased expression of NEFH and PCP4/PEP19 in the prefrontal cortex of alcoholics. Neurosci. Res. 2004;49:379–385. doi: 10.1016/j.neures.2004.04.002. [DOI] [PubMed] [Google Scholar]
- 76.Yan J, et al. A novel missense pathogenic variant in NEFH causing rare Charcot-Marie-Tooth neuropathy type 2CC. Neurol. Sci. 2021;42:757–763. doi: 10.1007/s10072-020-04595-z. [DOI] [PubMed] [Google Scholar]
- 77.Qiu Y, et al. Emerging views of OPTN (optineurin) function in the autophagic process associated with disease. Autophagy. 2022;18:73–85. doi: 10.1080/15548627.2021.1908722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wild P, et al. Phosphorylation of the autophagy receptor optineurin restricts Salmonella growth. Science. 2011;333:228–233. doi: 10.1126/science.1205405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ying H, Yue BY. Optineurin: The autophagy connection. Exp. Eye Res. 2016;144:73–80. doi: 10.1016/j.exer.2015.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Shen X, et al. Processing of optineurin in neuronal cells. J. Biol. Chem. 2011;286:3618–3629. doi: 10.1074/jbc.M110.175810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Yamano K, et al. Critical role of mitochondrial ubiquitination and the OPTN–ATG9A axis in mitophagy. J. Cell Biol. 2020;219:12144. doi: 10.1083/jcb.201912144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Sawa-Makarska J, et al. Reconstitution of autophagosome nucleation defines Atg9 vesicles as seeds for membrane formation. Science. 2020;369:eaaz7714. doi: 10.1126/science.aaz7714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Pensato V, et al. Sorting rare ALS genetic variants by targeted re-sequencing panel in Italian patients: OPTN, VCP, and SQSTM1 variants account for 3% of rare genetic forms. J. Clin. Med. 2020;9:412. doi: 10.3390/jcm9020412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Canosa A, et al. A familial amyotrophic lateral sclerosis pedigree discordant for a novel p. Glu46Asp heterozygous OPTN variant and the p. Ala5Val heterozygous SOD1 missense mutation. J. Clin. Neurosci. 2020;75:223–225. doi: 10.1016/j.jocn.2020.03.032. [DOI] [PubMed] [Google Scholar]
- 85.Mou Y, et al. OPTN variants in ALS cases: A case report of a novel mutation and literature review. Neurol. Sci. 2022;43:5391–5396. doi: 10.1007/s10072-022-06125-5. [DOI] [PubMed] [Google Scholar]
- 86.Evans CS, Holzbaur EL. Lysosomal degradation of depolarized mitochondria is rate-limiting in OPTN-dependent neuronal mitophagy. Autophagy. 2020;16:962–964. doi: 10.1080/15548627.2020.1734330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Medchalmi S, Tare P, Sayyad Z, Swarup G. A glaucoma-and ALS-associated mutant of OPTN induces neuronal cell death dependent on Tbk1 activity, autophagy and ER stress. FEBS J. 2021;288:4576–4595. doi: 10.1111/febs.15752. [DOI] [PubMed] [Google Scholar]
- 88.Feldman, M., Benatar, M., Lavi, E. S., Latchman, K. & Singer, C. (AAN Enterprises, 2023).
- 89.Wang C, et al. Isoforsythiaside attenuates Alzheimer’s disease via regulating mitochondrial function through the PI3K/AKT pathway. Int. J. Mol. Sci. 2020;21:5687. doi: 10.3390/ijms21165687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Xiao Q, et al. AMP-activated protein kinase-dependent autophagy mediated the protective effect of sonic hedgehog pathway on oxygen glucose deprivation-induced injury of cardiomyocytes. Biochem. Biophys. Res. Commun. 2015;457:419–425. doi: 10.1016/j.bbrc.2015.01.006. [DOI] [PubMed] [Google Scholar]
- 91.Yin X, et al. Downregulated AEG-1 together with inhibited PI3K/Akt pathway is associated with reduced viability of motor neurons in an ALS model. Mol. Cell. Neurosci. 2015;68:303–313. doi: 10.1016/j.mcn.2015.08.009. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data in this study are included in the supplementary data. The raw data used for machine learning is shown in Supplementary File 3, and the first two rows (file descriptions) must be removed before use. The raw counts of miRNA are also available from the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO) database of the accession number GSE168714.
The machine learning platform RapidMiner Studio is available at https://rapidminer.com/. The process is shown in Supplementary File 2, and the parameters are in the “Methods” section.