

Key Points
ABT-199, a new Bcl-2–specific BH3 mimetic, is efficacious in vivo against mouse lymphomas without provoking thrombocytopenia.
Treating mouse lymphomas with BH3 mimetic ABT-737 combined with bortezomib or purvalanol achieved long-term remission.
Abstract
BH3-only proteins trigger the stress apoptosis pathway and chemical mimetics have great potential for cancer therapy. BH3-only proteins inhibit antiapoptotic members of the Bcl-2 family. Promising BH3 mimetic ABT-737 and the related orally available compound ABT-263 (navitoclax) bind avidly to antiapoptotic Bcl-2, Bcl-xL, and Bcl-w. However, their interaction with Bcl-xL provokes thrombocytopenia, which has proven to be the dose-limiting toxicity. We have tested the efficacy of ABT-199, a new Bcl-2–specific BH3 mimetic, against aggressive progenitor cell lymphomas derived from bitransgenic myc/bcl-2 mice. As a single agent, ABT-199 was as effective as ABT-737 in prolonging survival of immunocompetent tumor-bearing mice without causing thrombocytopenia. Both drugs acted rapidly but, contrary to prevailing models, their apoptotic activity did not rely upon the BH3-only protein Bim. When ABT-737 was combined with the proteosome inhibitor bortezomib or CDK inhibitor purvalanol, many treated animals achieved long-term remission.
Introduction
Whether cells live or die by apoptosis when confronted with diverse stresses, including radiation and chemotherapy, is determined by interactions between opposing factions of the Bcl-2 family of proteins.1 Bcl-2 prevents apoptosis, as do Bcl-xL, Bcl-w, Mcl-1, and A1, but other close relatives Bax and Bak instead provoke apoptosis. Stress signals induce distant relatives known as BH3-only proteins, which bind avidly to a hydrophobic groove on prosurvival proteins, preventing them from restraining any activated Bax or Bak molecules. Certain BH3-only proteins (Bim, cleaved Bid, and perhaps Puma, Noxa) can also bind weakly and transiently to Bax and/or Bak, triggering their conformational change and homo-oligomerization on the outer mitochondrial membrane. As a consequence, cytochrome c is released into the cytoplasm and provokes activation of the proteases (caspases) that demolish the cell.
Chemical mimetics of BH3-only proteins represent an exciting new class of cancer therapeutic. The most promising thus far have been ABT-737 and the related orally available compound ABT-263 (navitoclax), which bind avidly to Bcl-2, Bcl-xL, and Bcl-w but not to Mcl-1 or A1.2,3 Both have significant efficacy against human tumor cell lines having high levels of Bcl-2 or Bcl-xL but little Mcl-1,4,5 particularly lymphoid malignancies and small cell lung carcinoma.2,3,6 However, because Bcl-xL is critical for the survival of platelets,7,8 ABT-737 and ABT-263 induced transient thrombocytopenia in preclinical trials, and the dose-limiting toxicity for navitoclax proved to be thrombocytopenia.9,10
Abbott Laboratories has recently developed a high-affinity Bcl-2–selective BH3 mimetic, ABT-199, which spared human platelets in vitro and dog platelets in vivo.11 Tumor regression was achieved for xenografts of human lymphoma cell lines and, excitingly, the first clinical tests for refractory chronic lymphocytic leukemia resulted in rapid tumor lysis in 3 of 3 patients.11
We have investigated the efficacy of ABT-199 in a preclinical model responsive to ABT-737: immunocompetent mice transplanted with lymphomyeloid progenitor cell tumors derived from Eμ-myc/Eμ-bcl-2 (hereafter myc/bcl-2) transgenic mice.12 These lymphomas have high levels of Bim, which is thought to be critical for responsiveness.13 To test the requirement for Bim, we also generated and treated bim−/− myc/bcl-2 tumors. In addition, we trialed combination therapy with ABT-737 and either the proteosome inhibitor bortezomib or the cyclin-dependent kinase (CDK) inhibitor purvalanol.
Study design
Nonirradiated C57BL/6 mice were injected (intravenously) with 3 × 106 lymphoma cells (3 mice per treatment arm per tumor) and monitored by tail bleed. Starting on day 11, when the white blood cell (WBC) count had become elevated, the BH3 mimetic or relevant vehicle was administered daily for 10 days (ABT-737: Abbott Laboratories (Chicago, IL), 75 mg/kg intraperitoneally in 30% propylene glycol, 5% Tween 80, 10% dimethylsulfoxide, 3.3% dextrose in water, pH 4; ABT-199: Abbott Laboratories, 100 mg/kg, oral gavage in 60% phosal 50PG (standardized phosphatidylcholine concentrate with at least 50% PC and propylene glycol; Phospholipid GmbH, Cologne, Germany), 30% PEG (polyethylene glycol) 400, 10% ethanol).
Results and discussion
All procedures involving animals were approved by the Walter and Eliza Hall Institute Animal Ethics Committee. Mice were transplanted with bim+/+ or bim−/− myc/bcl-2 lymphoma cells, and treatment with ABT-737 or ABT-199 was initiated once WBC counts were elevated (day 11). The impact of ABT-737 and ABT-199 on tumor-bearing mice was rapid, with the WBC count dropping within 3 hours (Figure 1A-B). By day 6, the WBC count had dropped further in most mice, except for 1 bim−/− lymphoma (no. 324), which had a mixed phenotype (see Figure 1 legend; supplemental Figure 1A, available on the Blood website).
Figure 1.

Comparison of responsiveness to ABT-737 and ABT-199. (A-B) WBC counts, (C-D) platelet counts, and (E-F) Kaplan-Meier survival curves of mice transplanted with 4 bim+/+ (nos. 9, 12, 16, and 47) and 3 bim−/− (nos. 58, 96, and 324) myc/bcl-2 lymphomas and treated with ABT-737, ABT-199, or the respective vehicles (3 mice per treatment arm for each independent tumor). Blood analyses were performed at 0 hours, 3 hours, and 6 days after starting treatment (day 11 after transplantation), using an ADVIA 2120 hematology analyzer (Siemens Australia New Zealand, VIC, Australia). Bars represent mean + SEM; significant changes observed at 3 hours are indicated: *P < .05, **P < .01, ***P < .001, Student t test. The x-axis in panels E and F indicates days elapsed since start of treatment; the bar indicates the duration of treatment (10 days). Significance for Kaplan-Meier survival curves was determined using the log-rank (Mantel-Cox) test. The median survival of bim+/+ lymphomas treated with ABT-737 was 21.5 days vs 14 days for vehicle (P = .0088), and 23.5 days with ABT-199 vs 13 days for vehicle (P = .0003). The median survival of bim−/− lymphomas treated with ABT-737 was 20.5 days vs 14 days for vehicle (P < .0001), and 19.5 days with ABT-199 vs 13 days for vehicle (P < .0001). One bim−/− lymphoma (no. 324) rebounded early following treatment; unusually, it comprised 50% progenitor (B220+CD4+) and 50% B-lymphoid (B220+CD4−) cells, the latter having higher levels of Mcl-1 (see supplemental Figure 1A), which would confer greater resistance.
Platelet counts were low prior to treatment, presumably due to the tumor burden (Figure 1C-D). Following the first dose of ABT-737, platelet counts dropped significantly within 3 hours but had rebounded by day 6, as previously reported.7 In mice treated with ABT-199, however, there was no drop in platelets at 3 hours and by day 6 they had increased to approximately normal levels, most likely due to the reduction in tumor cells. Thus, as anticipated for a Bcl-2–selective inhibitor, ABT-199 treatment did not cause acute thrombocytopenia.
Mice treated with the BH3 mimetics lived significantly longer than controls (Figure 1E-F). Those transplanted with bim+/+ lymphomas responded equally well to ABT-737 and ABT-199, their median survival being extended by approximately 10 days (Figure 1E). Indeed, had the treatment been continued, the mice may well have survived longer.
ABT-737 has been thought to act by displacing Bim from Bcl-2, thereby making it available to neutralize Mcl-1 or A1, which would otherwise contribute to resistance.13 Surprisingly, however, the bim−/− tumors responded well to treatment. With ABT-199, survival was only modestly less than for those with bim+/+ tumors (P = .039) and with ABT-737, the response was comparable (Figure 1E-F). These results indicate that Bim is not essential for apoptosis induced by these BH3 mimetics, at least in this tumor type.
We hypothesized that bim−/− tumors might have had a compensatory increase in another BH3-only protein. However, the level of Bid, Noxa, and Bad did not vary between tumors and, while some had higher Puma or Bmf than others, there was no consistent difference between the bim−/− and bim+/+ tumors (supplemental Figure 1B). Presumably, a combination of other BH3-only proteins had sufficed to replace Bim. It will be important to determine whether these findings extend to other Bcl-2–dependent tumor types, particularly in a clinical setting.
Long-term exposure to ABT-737 in vitro can result in the emergence of resistant clones through upregulation of antiapoptotic Bcl-2–related proteins.14 To test whether selection was occurring during treatment in vivo, we compared 5 tumors from sick mice that had been treated earlier with a BH3 mimetic or vehicle (supplemental Figure 2). No increase in endogenous Bcl-xL, Mcl-1, or Bcl-2 was observed but transgenic Bcl-2 was higher in certain treated tumors (bim+/+, no. 16 and, to a lesser extent, bim+/+, no. 9). Whether treatment selects those cells having higher Bcl-2 or induces higher expression is unclear. Of note, the duration of treatment used here (10 days) is comparatively short relative to expected clinical regimens.
Despite their encouraging response, all mice treated solely with a BH3 mimetic eventually died of their tumors (within 30 days following cessation of treatment). We previously showed striking synergy between ABT-737 and low-dose cyclophosphamide, achieving many complete remissions for mice transplanted with myc/bcl-2 lymphomas, and attributed the synergy to neutralization of Mcl-1 by BH3-only proteins Noxa and Puma induced by cyclophosphamide (via p53).12 We have now tested ABT-737 in combination with 2 agents expected to lower Mcl-1 levels: the proteasome inhibitor bortezomib and CDK (1, 2, 4) inhibitor purvalanol A (Figure 2; supplemental Tables 1-2). Although proteasome inhibition can lead to accumulation of Mcl-1,15 it also results in an increase in Noxa (eg, Gomez-Bougie et al16). CDK inhibitors downregulate Mcl-1 transcription17 and purvalanol A has been shown to induce apoptosis of Myc-overexpressing cells and extend survival of mice transplanted with Eμ-myc tumors.18 For direct comparison with the earlier study, treatment was started on day 4 rather than on day 11. None of the lymphomas responded to bortezomib or purvalanol A as single agents. Tumor no. 9 responded better to ABT-737 plus bortezomib than ABT-737 alone (P = .02), the response being comparable (P = .19) to that achieved with ABT-737 and low-dose cyclophosphamide, but ABT-737/purvalanol was not significantly better than ABT-737 alone (P = .12). For tumor no. 12, there was no significant benefit with ABT-737 plus bortezomib versus ABT-737 alone (P = .52), but the outcome with ABT-737/purvalanol was comparable with that achieved with ABT-737/cyclophosphamide (P = .47). For tumor no. 16, ABT-737 produced complete remission as a single agent, with all mice remaining healthy until the experiment was terminated at 150 days (supplemental Table 2); the greater sensitivity may be due to higher levels of Puma and Noxa in this tumor (supplemental Figure 1B). When treated with ABT-737 from day 11 rather than day 4, however, all mice transplanted with tumor no. 16 had died by day 50 (Figure 1E). This dramatic difference in outcome most likely reflects the relative tumor burden at the outset of treatment and longer treatment may have been more effective.
Figure 2.
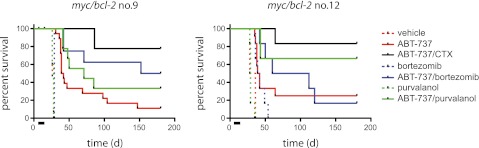
Combination therapy with ABT-737 extends survival. Safe regimens were established from previously published data12,18,19 and our own dose-finding studies (data not shown). Nonirradiated C57BL/6 recipients were transplanted with 3 independent bim+/+ myc/bcl-2 lymphomas (nos. 9, 12, 16) (3 × 106 tumor cells per mouse) 4 days prior to commencement of treatment. ABT-737 was administered intraperitoneally at 75 mg/kg per day for 10 days (bar), starting on day 1; cyclophosphamide (CTX; 50 mg/kg) intraperitoneally on days 3, 8; bortezomib (0.5 mg/kg) intraperitoneally on days 1, 3, 5, 8, 10; purvalanol A (20 mg/kg) intraperitoneally on days 1, 3, 5, 8, 10, alone or in combination with ABT-737, with at least 6 mice per indicated treatment arm (supplemental Tables 1-2). With tumor 16, all mice treated with ABT-737 alone remained healthy until the experiment was terminated at day 150 (supplemental Table 2 and data not shown; controls treated with vehicle were all dead by day 35).
Early clinical trials for treating CLL with navitoclax9 and now ABT-19911 show promising potential. In view of our results, combination therapy with cyclophosphamide, bortezomib (velcade), and clinically approved CDK inhibitors would be worth considering. Others have also reported encouraging in vivo synergies with ABT-7376,19 and, very recently, with ABT-199.11
Supplementary Material
Acknowledgments
The authors thank Abbott Laboratories for generously providing ABT-737 and ABT-199; Kylie D. Mason, David C. S. Huang, Clare Scott, and Andrew W. Roberts for useful discussions; G. Siciliano, K. McKenzie, and T. Camilleri for excellent technical assistance; and A. Strasser for reagents.
This work was supported by National Health and Medical Research Council (NHMRC) (Australia) program grant 461221, a NHMRC Career Development Fellowship (C.J.V.), National Institutes of Health National Cancer Institute (NCI) grant CA43540, Leukemia & Lymphoma Society Specialized Center for Research grant 7015-02, the Australian Cancer Research Foundation (Centre for Therapeutic Target Drug Discovery), and infrastructure support to The Walter and Eliza Hall Institute (WEHI) from the NHMRC Independent Research Institute Infrastructure Support Scheme (IRIISS) and the Victorian State Government Operational Infrastructure Support (OIS).
Footnotes
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Authorship
Contribution: C.J.V. and S.C. conceived the studies, planned the experiments, analyzed data, and wrote the manuscript; and C.J.V. performed the experiments.
Conflict-of-interest disclosure: The authors declare no competing financial interests. The Walter and Eliza Hall Institute had a tripartite research collaboration with Genentech Inc and Abbott Laboratories at the time of these studies.
Correspondence: Suzanne Cory, Molecular Genetics of Cancer Division, The Walter and Eliza Hall Institute of Medical Research, 1G Royal Parade, Parkville 3052, VIC, Australia; e-mail: cory@wehi.edu.au.
References
- 1.Strasser A, Cory S, Adams JM. Deciphering the rules of programmed cell death to improve therapy of cancer and other diseases. EMBO J. 2011;30(18):3667–3683. doi: 10.1038/emboj.2011.307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Oltersdorf T, Elmore SW, Shoemaker AR, et al. An inhibitor of Bcl-2 family proteins induces regression of solid tumours. Nature. 2005;435(7042):677–681. doi: 10.1038/nature03579. [DOI] [PubMed] [Google Scholar]
- 3.Tse C, Shoemaker AR, Adickes J, et al. ABT-263: a potent and orally bioavailable Bcl-2 family inhibitor. Cancer Res. 2008;68(9):3421–3428. doi: 10.1158/0008-5472.CAN-07-5836. [DOI] [PubMed] [Google Scholar]
- 4.van Delft MF, Wei AH, Mason KD, et al. The BH3 mimetic ABT-737 targets selective Bcl-2 proteins and efficiently induces apoptosis via Bak/Bax if Mcl-1 is neutralized. Cancer Cell. 2006;10(5):389–399. doi: 10.1016/j.ccr.2006.08.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Konopleva M, Contractor R, Tsao T, et al. Mechanisms of apoptosis sensitivity and resistance to the BH3 mimetic ABT-737 in acute myeloid leukemia. Cancer Cell. 2006;10(5):375–388. doi: 10.1016/j.ccr.2006.10.006. [DOI] [PubMed] [Google Scholar]
- 6.Ackler S, Mitten MJ, Foster K, et al. The Bcl-2 inhibitor ABT-263 enhances the response of multiple chemotherapeutic regimens in hematologic tumors in vivo. Cancer Chemother Pharmacol. 2010;66(5):869–880. doi: 10.1007/s00280-009-1232-1. [DOI] [PubMed] [Google Scholar]
- 7.Mason KD, Carpinelli MR, Fletcher JI, et al. Programmed anuclear cell death delimits platelet life span. Cell. 2007;128(6):1173–1186. doi: 10.1016/j.cell.2007.01.037. [DOI] [PubMed] [Google Scholar]
- 8.Zhang H, Nimmer PM, Tahir SK, et al. Bcl-2 family proteins are essential for platelet survival. Cell Death Differ. 2007;14(5):943–951. doi: 10.1038/sj.cdd.4402081. [DOI] [PubMed] [Google Scholar]
- 9.Roberts AW, Seymour JF, Brown JR, et al. Substantial susceptibility of chronic lymphocytic leukemia to BCL2 inhibition: results of a phase I study of navitoclax in patients with relapsed or refractory disease. J Clin Oncol. 2012;30(5):488–496. doi: 10.1200/JCO.2011.34.7898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wilson WH, O’Connor OA, Czuczman MS, et al. Navitoclax, a targeted high-affinity inhibitor of BCL-2, in lymphoid malignancies: a phase 1 dose-escalation study of safety, pharmacokinetics, pharmacodynamics, and antitumour activity. Lancet Oncol. 2010;11(12):1149–1159. doi: 10.1016/S1470-2045(10)70261-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Souers AJ, Leverson JD, Boghaert ER, et al. ABT-199, a potent and selective BCL-2 inhibitor, achieves antitumor activity while sparing platelets. Nat Med. 2013;19(2):202–208. doi: 10.1038/nm.3048. [DOI] [PubMed] [Google Scholar]
- 12.Mason KD, Vandenberg CJ, Scott CL, et al. In vivo efficacy of the Bcl-2 antagonist ABT-737 against aggressive Myc-driven lymphomas. Proc Natl Acad Sci U S A. 2008;105(46):17961–17966. doi: 10.1073/pnas.0809957105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Del Gaizo Moore V, Brown JR, Certo M, et al. Chronic lymphocytic leukemia requires BCL2 to sequester prodeath BIM, explaining sensitivity to BCL2 antagonist ABT-737. J Clin Invest. 2007;117(1):112–121. doi: 10.1172/JCI28281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Yecies D, Carlson NE, Deng J, et al. Acquired resistance to ABT-737 in lymphoma cells that up-regulate MCL-1 and BFL-1. Blood. 2010;115(16):3304–3313. doi: 10.1182/blood-2009-07-233304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nencioni A, Hua F, Dillon CP, et al. Evidence for a protective role of Mcl-1 in proteasome inhibitor-induced apoptosis. Blood. 2005;105(8):3255–3262. doi: 10.1182/blood-2004-10-3984. [DOI] [PubMed] [Google Scholar]
- 16.Gomez-Bougie P, Wuillème-Toumi S, Ménoret E, et al. Noxa up-regulation and Mcl-1 cleavage are associated to apoptosis induction by bortezomib in multiple myeloma. Cancer Res. 2007;67(11):5418–5424. doi: 10.1158/0008-5472.CAN-06-4322. [DOI] [PubMed] [Google Scholar]
- 17.Raje N, Kumar S, Hideshima T, et al. Seliciclib (CYC202 or R-roscovitine), a small-molecule cyclin-dependent kinase inhibitor, mediates activity via down-regulation of Mcl-1 in multiple myeloma. Blood. 2005;106(3):1042–1047. doi: 10.1182/blood-2005-01-0320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Goga A, Yang D, Tward AD, et al. Inhibition of CDK1 as a potential therapy for tumors over-expressing MYC. Nat Med. 2007;13(7):820–827. doi: 10.1038/nm1606. [DOI] [PubMed] [Google Scholar]
- 19.Paoluzzi L, Gonen M, Bhagat G, et al. The BH3-only mimetic ABT-737 synergizes the antineoplastic activity of proteasome inhibitors in lymphoid malignancies. Blood. 2008;112(7):2906–2916. doi: 10.1182/blood-2007-12-130781. [DOI] [PubMed] [Google Scholar]
- 20.Adams JM, Harris AW, Pinkert CA, et al. The c-myc oncogene driven by immunoglobulin enhancers induces lymphoid malignancy in transgenic mice. Nature. 1985;318(6046):533–538. doi: 10.1038/318533a0. [DOI] [PubMed] [Google Scholar]
- 21.Strasser A, Whittingham S, Vaux DL, et al. Enforced BCL2 expression in B-lymphoid cells prolongs antibody responses and elicits autoimmune disease. Proc Natl Acad Sci U S A. 1991;88(19):8661–8665. doi: 10.1073/pnas.88.19.8661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bouillet P, Cory S, Zhang L-C, et al. Degenerative disorders caused by Bcl-2 deficiency prevented by loss of its BH3-only antagonist Bim. Dev Cell. 2001;1(5):645–653. doi: 10.1016/s1534-5807(01)00083-1. [DOI] [PubMed] [Google Scholar]
- 23.Strasser A, Harris AW, Bath ML, et al. Novel primitive lymphoid tumours induced in transgenic mice by cooperation between myc and bcl-2. Nature. 1990;348(6299):331–333. doi: 10.1038/348331a0. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.