

Abstract
Toll-like receptors (TLRs) are crucial components of innate immunity that participate in host defense against microbial pathogens. We evaluated the expression and function of TLRs in human retinal pigment epithelial (RPE) cells. Real time PCR analysis revealed gene expression for TLRs 1–7, 9, and 10 in RPE cells. TLRs 1 and 3 were the most highly expressed TLRs. Protein expression for TLRs 2, 3, and 4 was observed on RPE cells and this expression was augmented by treatment with poly I:C or interferon-γ (IFN-γ). TLR 3 is the receptor for dsRNA, an intermediate of virus replication. Because RPE cells express TLR 3 and are frequently the site of virus replication within the retina, we evaluated TLR 3 signaling. RPE cells treated with poly I:C produced IFN-β but not IFN-α, and this was inhibited by the treatment of RPE cells with anti-TLR 3 antibody. Human recombinant IFN-β was shown to be biologically active on RPE cells by inhibiting viral replication. Poly I:C treatment of RPE resulted in an increase in the production of IL-6, IL-8, MCP-1, and sICAM-1. The presence of TLRs on RPE cells and the resultant TLR signaling in RPE cells suggest that these molecules may play an important role in innate and adaptive immune responses within the retina.
Keywords: Toll-like receptors, dsRNA, Interferon-β, Retinal pigment epithelial cells, Innate immunity, Cytokines, Poly I:C
1. Introduction
Toll-like receptors, TLRs, are a family of evolutionary conserved innate immune recognition molecules that recognize molecular patterns associated with microbial pathogens (Medzhitov and Janeway, 1997). They constitute a first line of defense against a variety of pathogens and play a critical role in initiating the innate immune response. TLR recognition of these specific microbial patterns leads to a signal transduction cascade that generates a rapid and robust inflammatory response marked by cellular activation and cytokine release Gordon, 2002, Schnare et al., 2001.
To date, 10 mammalian TLRs have been identified and each receptor appears to be involved in the recognition of a unique set of microbial patterns Gordon, 2002, Zuany-Amorim et al., 2002. For example, TLR 2 recognizes various ligands expressed by Gram-positive bacteria, whereas TLR 3 engages dsRNA and TLR 4 is specific for Gram-negative bacteria lipopolysaccharides (LPS; Alexopoulou et al., 2001, Campos et al., 2001, Johnson et al., 2002, Opitz et al., 2001). TLR 5, on the other hand, recognizes bacterial flagellin, while TLRs 7 and 8 interact with antiviral compounds and TLR 9 binds bacterial DNA Bauer et al., 2001, Gewirtz et al., 2001, Ito et al., 2002. Recently, TLRs were observed to influence the development of the adaptive immune response presumably through the activation of antigen-presenting cells (APC; Schjetne et al., 2003). The most compelling evidence comes from studies on dendritic cells (Banchereau et al., 2003). It has previously been shown that multiple TLRs are found on dendritic cells. Signaling through these receptors augments antigen presentation by driving cell maturation, up-regulating expression of costimulatory molecules on the cell, and inducing cytokines (Palucka and Banchereau, 2002). Thus, TLRs may serve as a unique link between innate and adaptive immunity.
The retinal pigment epithelium consists of a single layer of cells of neural ectoderm origin, which lie between the photoreceptors of the neural retina and the blood-rich choroid. These vitally important cells phagocytize the shed discs from the photoreceptor outer segments, recycling their components, such as retinoids. Additional functions of this monolayer of cells include the transport of nutrients from the choroid into the retina and the transport of waste in the reverse direction. This cell also adsorbs light and provides adhesive properties for the retina Bok, 1985, Bok, 1993.
The retinal pigment epithelial (RPE) cell also plays a key role in a variety of retinal pathologic processes. Inherited retinal degenerative diseases can be associated with mutations of RPE cellular genes Hamel et al., 1993, Morimura et al., 1998, Morimura et al., 1999. Moreover, degenerative diseases, such as age-related macular degeneration and diabetic retinopathy, can be associated with early damage to the RPE cell Cai et al., 2000, Lutty et al., 1999. In addition, a variety of in vivo and in vitro studies have identified this cell as an ideal target for infectious agents such as cytomegalovirus (CMV), Toxoplasma gondii and coronavirus Detrick et al., 1996, Detrick et al., 2001, Hooks et al., 1993, Nagineni et al., 2000, Nagineni et al., 2002. Furthermore, this cell is a rich source of cytokines, chemokines, and growth factors that may contribute to or limit pathologic processes Chin et al., 2001, Momma et al., 2003. Recently, the RPE cell has been shown to play a pivotal role in the immune system. It has been demonstrated that interferon-γ (IFN-γ) treatment up-regulates the expression of both MHC class-I and -II molecules on RPE cells (Percopo et al., 1990). Moreover, RPE cells have been reported to act as APC in the retina (Percopo et al., 1990). Hence, RPE cells can incorporate pathogens, produce a variety of cytokines, and present pathogen-derived peptides to sensitized T cells. This finding, that RPE cells function as APC in the retina, extends the activities of this cell beyond its participation as a first line defense cell and underscores its important role in adaptive immunity. Therefore, based on these observations, it was of interest to further define the role of this epithelial cell in innate and adaptive immune responses within the retina.
To date, there is no information about the presence of TLRs within the retina. The results of this study demonstrate that RPE cells do indeed constitutively express distinct types of TLRs and that their expressions are modulated in the presence of dsRNA and cytokines. TLR 3 is a receptor for dsRNA, and dsRNA is a common replication intermediate for many viruses. Moreover, TLR 3 has been described as a specific TLR because it displays the most restricted cellular expression pattern (Janssens and Beyaert, 2003). Because TLR 3 gene expression was identified in RPE cells and because the RPE cells are a site of replication for both RNA and DNA viruses, we further investigated TLR 3 signaling in these cells. Our data suggest that the binding of poly I:C, an analog of dsRNA, to TLR 3 on human RPE cells resulted in the production of IFN-β and other cytokines, chemokines, and adhesion molecules. Thus, TLR 3 signaling within the retina may provide additional protective molecules to mediate viral infections.
2. Materials and methods
2.1. Cells and reagents
Affinity purified, monoclonal, antihuman TLR 2 and TLR 3 antibodies were purchased from Imgenex (San Diego, CA) while antihuman TLR 4 (HTA 125) antibodies were purchased from eBiosciences (San Diego, CA). Polyclonal antibodies to TLR 3 were obtained from Santa Cruz Biotechnology (Santa Cruz, CA), LPS (Salmonella typhosa), poly I:C, and poly dI:dC were purchased from Sigma (St Louis, MO). The recombinant human (IFN-γ) and IFN-β were procured from Roche Molecular Biochemicals (Indianapolis, IN). RNA STAT-60 was obtained from Tel-Test (Friendswood, TX). Minimal essential media (MEM), fetal bovine serum (FBS), penicillin/streptomycin/fungizone, nonessential amino acids, and normal horse serum (NHS) were purchased from Life Technologies/Gibco (Gaithersburg, MD). GeneAmp RNA PCR kits and Taqman reagents were obtained from Perkin Elmer (Branchburg, NJ).
Human RPE cell cultures were prepared from donor eyes and grown in MEM supplemented with 10% FBS, nonessential amino acids, and penicillin/streptomycin/fungizone in a 5% CO2, humidified 37 °C incubator. Characterization of these cells has been described previously Li et al., 1999, Nagineni et al., 1994, Nagineni et al., 1996 Briefly, these cells demonstrated a hexagonal morphology when grown to confluence and formed monolayers with distinct intracellular boundaries. Homogeneity of the cultures was established by positive immunostaining with monoclonal antibodies to cytokeratin, an epithelial cell-specific cytoskeletol protein. For the experiments described in this paper, human RPE cultures, at passages 7 to 12, were used. A human monocyte cell line, U937, was grown in an RPMI-1640 medium supplemented with 10% FBS (ATCC, Manassas, VA).
2.2. Analysis of TLR gene expression in human RPE by RT-PCR
Total RNA prepared from confluent monolayers of human RPE cells and from suspension cultures of U937 was used to evaluate the constitutive expression of TLR mRNA. To study the effects of TLR activators, human RPE cells were washed with serum-free media (SFM) and incubated in SFM for 12 h in the presence of poly I:C (100 μg/ml ), LPS (5 μg/ml), or IFN-γ (100 U/ml). Total RNA was prepared from the cell cultures by using the RNA STAT-60 extraction solution. One μg of total RNA was used for each RT-PCR reaction. The RT-PCR procedure was performed using an RNA PCR kit (Perkin-Elmer) according to the manufacturer's instructions. PCR products were separated by gel electrophoresis, photographed under UV light, and integrated with an image acquisition system (Eagle Eye, Stratagene, San Diego, CA). The following primer pairs were used for the analysis of TLRs, costimulator molecules, and GAPDH by RT-PCR: TLR 1 (219 bp) 5′-CTATACACCAAGTTGTCAGC-3′ and 5′-GTCTCCAACTCAGTAAGGTG-3′; TLR 2 (346 bp) 5′-GCCAAAGTCTTGATTGATTGG-3′ and 5′-TTGAAGTTCTCCAGCTCCTG-3′; TLR 3 (304 bp) 5′-GATCTGTCTCATAATGGCTTG-3′ and 5′-GACAGATTCCGAATGCTTGTG-3′; TLR 4 (506 bp) 5′-TGGATACGTTTCCTTATAAG-3′ and 5′-GAAATGGAGGCACCCCTTC-3′; TLR 5 (437 bp) 5′-TAGCTCCTAATCTGATG-3′ and 5′-CCATGTGAAGTCTTTGCT GC-3′; TLR 7 (388 bp) 5′-TCTACCTGGGCCAAAACT GTT-3 and 5′-GGCACATGCTGAAGAGAGTTA-3'; TLR 8 (443 bp) 5′-GCCAGCGAGTCTCACTGAACT-3′ and 5′-GCCAGGGCAGCCAACATA-3′; TLR 9 (259 bp) 5′-GTCCCCACTTCTCCATG-3′ and 5′-GGCACAGTCATGATGTTGTTG-3′; CD14 (426 bp) 5′-CTCAAC CTAGAGCCGTTTCT-3′ and 5′-CAGGATTGTCAGACAGGTCT-3′; MD2 (359 bp) 5′-TATTGGGTCTGCAACTCA T-3′ and 5′-CTCCCAGAAATAGCTTCAAC-3′; GAPDH (600 bp) 5′-CCACCCATGGCAAATTCCATGGCA-3′ and 5′-TCTAGACGGCAGGTCAGGTCCACC-3′ Faure et al., 2000, Ito et al., 2002, Jarrossay et al., 2001, Bauer et al., 2001, Tabeta et al., 2000.
2.3. Real time (Taqman) analysis of TLR gene expression
Total RNA was prepared from quiescent RPE and U937 by using an RNA STAT-60 reagent. Quantitative RT-PCR analysis of TLR in RPE and U937 was performed on an ABI Prism 7700 (Applied Biosystems, Foster City, CA) by using Taqman master mix reagent kits according to the manufacturer's instructions. FAM-labeled Taqman probes and primers for human GAPDH and Toll-like receptors 1–10 (Assays-on-Demand gene expression products) were obtained from Applied Biosystems. Standard curves were generated to GAPDH and TLRs 1 to 10 by ten-fold serial dilutions of RPE and/or U937 RNA. RNA samples were analyzed in triplicate under similar conditions as those of the standards in the same 96-well plates for 40 cycles. Fluorescence intensities obtained for the samples were used to calculate relative fluorescence units by normalizing to GAPDH fluorescence intensities. Results are expressed as relative fluorescence units of TLRs 1–10 mRNA levels in RPE and U937 cells.
2.4. Immunofluorescent staining of RPE cells for TLRs 2, 3, and 4
The RPE cells were seeded onto Lab-Tek tissue culture chamber slides (Nalge Nunc International, Naperville, IL). After 24 h, the cells were washed with SFM and stimulated with media, poly I:C (100 μg/ml), LPS (S. typhosa, 5 μg/ml), or with IFN-γ (100 U/ml) for 24 h. The slides were then fixed in equal parts of acetone/methanol and stored at −20 °C until analyzed. The slides were washed twice with PBS for 5 min and then treated with 10% NHS in PBS for 1 h at room temperature. The slides were overlaid with primary mouse antihuman TLRs 2, 3, or 4 monoclonal antibodies (20 μg/ml in PBS with 10% NHS) or with mouse IgG (control) and incubated for 1-h followed by five washes in PBS containing 1% NHS. Cells were then incubated for 1 h with biotin-labeled horse antimouse IgG (H+L; Vector Laboratories, Burlingame, CA). The cells were washed again five times as described previously. FITC-labeled streptavidin (20 μg/ml) was added and the cells were incubated for 30 min in the dark (Vector Laboratories). The slides were then washed twice, mounted, and examined with a fluorescent microscope.
2.5. EIA assays for cytokines, chemokines, and adhesion molecules
RPE cultures were grown to confluence in 24-well dishes. Cultures were washed with SFM and incubated in the presence of various concentrations of poly I:C or poly dI:dC for 24 h at 37 °C. Supernatants were collected and stored at −70 °C until analyzed. The concentration of IL-6, IL-8, MCP-1, sICAM-1, IFN-α, and IFN-β in the cell culture supernatant fluids was determined by EIA. The assay was performed according to manufacturer's instructions (Quantikine EIA kits, R&D Systems, Minneapolis, MN). The data were analyzed using the VERSAmax data analysis program (Molecular Devices, Sunnyvale, CA). Results from two representative experiments are presented as the means±S.D. of triplicate cytokine measurements.
2.6. Neutralization of poly I:C effects on RPE by TLR 3 antibody
RPE cultures were grown to confluence in 24-well plates in 10% FBS media. Media were removed and replaced with serum free media (SFM). After 4 h, media were removed and replaced with fresh SFM (1 ml per well) and polyclonal antibody to TLR 3 (10 ug IgG/ml). After a 1-h incubation at 37 °C, poly I:C was added to the wells to obtain a final concentration of 2 or 20 ug/ml. The cultures were further incubated for 24 h and culture supernatants were collected. The levels of secreted IFN-β were determined by EIA.
2.7. IFN bioassay: vesicular stomatitis virus plaque inhibition
RPE cultures were grown to confluence in 24-well plates. The cultures were washed and incubated for 20 h with serial ten-fold dilutions of recombinant human IFN-β or with media. The monolayers were washed and challenged with approximately 100 plaque-forming units (pfu) of vesicular stomatitis virus. One hour later, the virus inoculum was removed and the cells were washed and refed with 1 ml of MEM containing 0.75% methylcellulose and 2% FBS. After a 24-h incubation at 37 °C, the overlay medium was removed, the cells were fixed with ethanol and stained with Giemsa's solution, and the viral plaques were counted.
3. Results
3.1. Constitutive expression of TLRS on human RPE and human monocytes
Preliminary studies using RT-PCR analysis indicated that human RPE cells contained detectable amounts of mRNA for TLRs 1, 2, 3, 4, and 5. U937 cells are a human monocyte cell line that was used as a control. U937 cells contained detectable levels of mRNA for TLRs 1, 2, 4, 5, 7, and 9. TLR 3 mRNA was barely detected in the U937 cells. In contrast, TLR 3 mRNA was highly expressed in human RPE cells. Moreover, mRNA for two coreceptors for TLRs, CD14, and MD2 were detected in human RPE whereas the U937 cells expressed only CD14.
In order to more accurately define the constitutive expression of TLRs, we analysed mRNA obtained from the two cell types using real time RT-PCR analysis. As shown in Table 1 , real time RT-PCR analysis of RPE cells revealed the constitutive expression of varying levels of mRNA for TLRs 1, 2, 3, 4, 5, 6, 7, 9, and 10. TLR 8 was not detected and low levels of expression were noted for TLRs 2, 4, 7, and 9. TLRs 1 and 3 were the most highly expressed TLRs in RPE cells. When compared to U937 cells, RPE cells contained 42 times more mRNA for TLR 3.
Table 1.
Real Time PCR analyses of TLR mRNA expression in RPE and U937 cells
| TLR | Relative fluorescence intensities |
|||
|---|---|---|---|---|
| RPE cells | U937 cells | Fold differencea (RPE/U937) | ||
| TLR 1 | 81,280 | 17,541 | 5× (+) | |
| TLR 2 | 1968 | 8364 | 4× (−) | |
| TLR 3 | 26,897 | 644 | 42× (+) | |
| TLR 4 | 868 | 19,066 | 22× (−) | |
| TLR 5 | 24,303 | 7983 | 3× (+) | |
| TLR 6 | 21,268 | 20,210 | 1× (+) | |
| TLR 7 | 1336 | 12,939 | 10× (−) | |
| TLR 8 | 0 | 0 | 0 | |
| TLR 9 | 1211 | 13,856 | 11× (−) | |
| TLR 10 |
7700 |
10,915 |
1× (−) |
|
Real time RT-PCR was performed on an ABI Prism 7700. Standard curves were generated to GAPDH and TLRs 1–10 by ten-fold serial dilutions of RPE and/or U937 total RNA. RNA samples were analyzed in triplicate under similar conditions as those of the standards in the same 96-well plates for 40 cycles. Fluorescence intensities obtained for the samples were used to calculate relative fluorescence units by normalizing to GAPDH fluorescence intensities. Results are expressed as relative fluorescence units for TLRs 1–10 mRNA levels in RPE and U937 cells.
Fold difference in RPE cells relative to U937 cells: (+) fold increase in RPE cells; (−) fold decrease in RPE cells.
3.2. Modulation of TLR gene expression by poly I:C
We next wanted to determine if poly I:C (dsRNA) treatment altered TLR gene expression in RPE cells. The effect of poly I:C treatment on TLRs 2, 3, and 4 gene expressions in RPE cells is shown in Fig. 1 . NIH image analysis of this data indicated that poly I:C treatment increased gene expression of TLR 2 by 19%, TLR 3 by 59%, and TLR 4 by 68%. Gene expression for the coreceptors, CD14 and MD2, was also evaluated. Poly I:C treatment had no effect on CD14 and MD2.
Fig. 1.
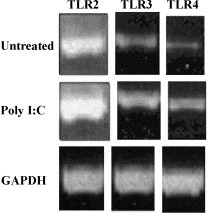
Modulation of TLR 2, TLR 3, and TLR 4 expression in human RPE cells by poly I:C. RPE cultures were grown to confluent monolayers in 100-mm dishes and incubated without (untreated) or with poly I:C (100 μg/ml) for 24 h. The procedure for total RNA preparation and RT-PCR is described in the Materials and methods section. RNA (1 μg) was used for each RT-PCR reaction. The sequences of the PCR primers and the expected sizes of PCR products are given in the Materials and methods section. All PCR reactions were subjected to 30 cycles. The results shown are from one representative experiment.
3.3. Detection of TLR protein on RPE cells by immunofluorescent staining
RPE cells propagated on culture slides were exposed to media alone or media containing poly I:C (100 μg/ml) or IFN-γ (100 U/ml; Fig. 2 ). After 24 h, the cells were washed and fixed with equal volumes of acetone and methanol and reacted with normal mouse IgG or with antibody directed to TLRs 2, 3, or 4. Staining was not observed in cells treated with normal mouse IgG (control). The intensity of TLR 2 reactivity was weak on untreated RPE cells, but this was enhanced by pretreatment with poly I:C or IFN-γ (data not shown). Intensity of TLR 3 reactivity was moderate in untreated RPE cells. This intensity was enhanced with poly I:C (Fig. 2A and B) or IFN-γ. Intensity of TLR 4 reactivity was moderate on untreated RPE cells but was augmented following pretreatment with IFN-γ (Fig. 2C and D) or with poly I:C. These data demonstrate that RPE cells express varying amounts of TLRs 2, 3, and 4 and that these receptors can be modulated by selected TLR activators. The pattern of staining for TLR 3 and TLR 4 in RPE cells was different. TLR 3 staining appears dispersed throughout the cytoplasm whereas TLR 4 staining appears to be more localized at the cell boarders.
Fig. 2.
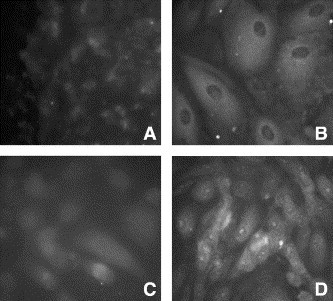
Immunofluorescent staining for TLR 3 and TLR 4 in RPE cells. Staining reactivity for TLR 3 is shown in untreated cells (A) and in cells treated for 24 h with poly I:C (100 μg/ml; B). Staining reactivity for TLR 4 is shown in untreated cells (C) and in cells treated for 24 h with IFN-γ (100 U/ml; D).
3.4. Induction of IFN-β by poly I:C-treated RPE cells
TLR 3 has only been detected on a limited number of cell types and recent studies indicate that TLR 3 is a receptor for dsRNA produced by viruses. Moreover, dsRNA binding to TLR 3 on the cell surface results in the production of type-1 IFNs. Therefore, we next investigated the possibility that treatment of RPE cells with synthetic dsRNA, poly I:C, would induce IFN. RPE cells were treated with varying concentrations of poly I:C or with poly dI:dC (50 mU/ml). Poly dI:dC is a synthetic double-stranded polydeoxyinosine/deoxycytosine (dsDNA) and is used as a negative control for poly I:C (dsRNA). The cells were incubated for 24 h and supernatant fluids were collected and assayed for IFN-α and IFN-β by EIA. As seen in Fig. 3A , poly I:C induced IFN-β in a dose-dependent manner. IFN-α was not detected in these samples or in untreated cells. Moreover, cells treated with poly dI:dC did not release IFN-α or IFN-β.
Fig. 3.
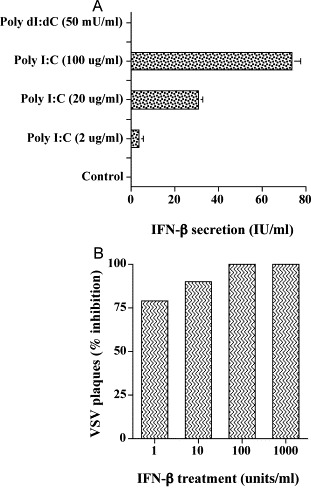
Secretion of IFN-β by human RPE cells (A) and antiviral activity of IFN-β (B). (A) Human RPE cell monolayers were grown to confluence. Cultures were washed with SFM twice and incubated in the presence of various concentrations of poly I:C or poly dI:dC for 24 h. The culture supernatants were collected and analyzed for IFN-α and -β by EIA. Results from three experiments conducted in duplicate are presented as the means±S.E. (B) Human RPE cultures were grown to confluence and were infected with Vesicular Stomatitis Virus (VSV) as described in the Materials and methods section. After 24 h, cultures were fixed and stained with Giemsa, and the plaques were counted. The data are presented as the means±S.E. of triplicate cultures obtained from a representative experiment.
Because we have shown that RPE cells can produce IFN-β, we next wanted to determine if IFN-β was biologically active on RPE cells. Cell cultures were treated with varying concentrations of recombinant human IFN-β. After incubation for 24 h, cells were challenged with approximately 100 pfu of VSV. Virus replication in RPE was evaluated by plaque assay. As seen in Fig. 3B, RPE cells were sensitive to the antiviral action of IFN-β. One unit of IFN-β inhibited VSV by 80%, whereas 10 and 100 units inhibited virus replication by 90 to 95%.
In order to evaluate the specificity of the poly I:C induction of IFN-β through TLR 3, we next performed antibody inhibition assays. RPE cells were pretreated with anti-TLR 3 antibody for 1 h and were then incubated with poly I:C for 24 h. Supernatant fluids were harvested and then analyzed for the presence of IFN-β. As seen in Fig. 4 , treatment with 2 or 20 ug of poly I:C induced 19.9±1.7 and 20.1±2.05 units of IFN-β, respectively. Pretreatment of the cells with anti-TLR 3 antibody reduced the levels of IFN-β produced to 3.7 and 0.17 (P<0.0001).
Fig. 4.
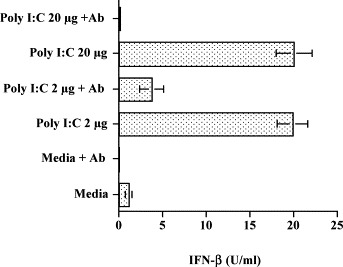
The effect of anti-TLR 3 antibody on poly I:C-induced IFN-β production in RPE cells. RPE cell cultures grown to confluence were washed with SFM and incubated with media or anti-TLR 3 antibody (10 ug/ml) for 1 h. Then, poly I:C was added to the appropriate wells to give a final concentration of 2 or 20 ug/ml. After 24 h of incubation, supernatant fluids were collected and the concentration of IFN-β was determined by EIA. Results presented were obtained from one representative experiment with quadruplicate samples.
3.5. Evaluation of poly I:C treatment of RPE cells: cytokines, chemokines, and adhesion molecules
We next investigated whether poly I:C treatment of RPE cells resulted in the modification of additional cellular functions such as the production of cytokines, chemokines, and adhesion molecules. RPE cells were incubated with varying concentrations of poly I:C for 48 h. The supernatant fluids were collected and evaluated by EIA for IL-6, IL-8, MCP-1, and sICAM-1 production. As seen in Fig. 5A–D , supernatant fluids from untreated RPE cells did not contain IL-6 or IL-8 and only very low levels of MCP-1 and sICAM-1. Poly I:C treatment of RPE cells resulted in a dose-dependent enhancement of IL-6, IL-8, MCP-1, and sICAM-1. Following poly I:C treatment at 2, 20, or 100 μg/ml, the concentration of IL-6 increased from 40 to 80 ng/ml, IL-8 increased from 9 to 29 ng/ml, MCP-1 increased from 50 to 160 ng/ml, and sICAM-1 increased from 2.5 to 8 ng/ml. In contrast, treatment of RPE cells under similar conditions with poly dI:dC did not enhance the secretion of IL-6, IL-8, MCP-1, and sICAM-1 (Fig. 5).
Fig. 5.
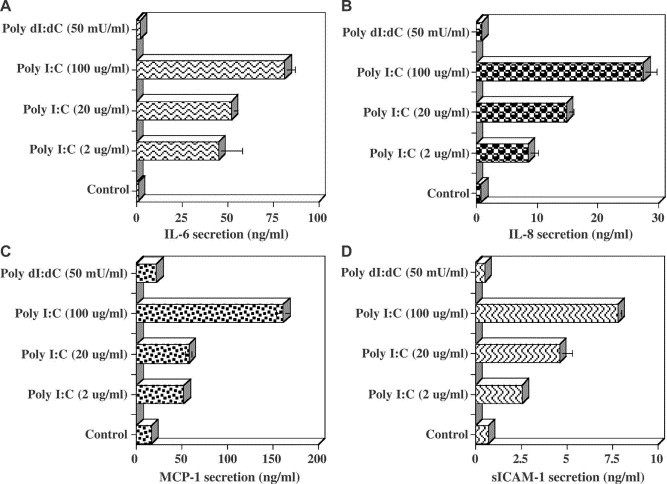
The effects of poly I:C treatment on the production of cytokines by human RPE cells. RPE cell cultures grown to confluence in 24-well plates were washed with SFM and incubated without (control) or with SFM containing various concentrations of poly I:C or poly dI:dC. After 24 h, the culture supernatants were collected and the concentrations of IL-6, IL-8, MCP-1, and sICAM-1 were determined by EIA. Results presented for IL-6 (A), IL-8 (B), MCP-1 (C), and ICAM-1 (D) were obtained from duplicate samples of two representative experiments.
4. Discussion
TLRs are critical elements in the host defense against microbial pathogens (Takeda et al., 2003). In this report, we demonstrate for the first time the presence of TLRs on human RPE cells. Real time PCR analysis of TLR gene expression identified TLRs 1–7, 9 and 10 in human RPE cells. Furthermore, human RPE cells highly expressed TLR 3. Protein expression for TLRs 2, 3 and 4 was also observed on RPE cells and this expression was augmented by treatment with poly I:C or IFN-γ. Because TLR 3 is found on a limited number of cells and was highly expressed in RPE cells, we performed studies to analyze signaling through TLR 3. The interaction of poly I:C with RPE cells resulted in the secretion of IFN-β as well as IL-6, IL-8, MCP-1, and sICAM-1. Moreover, we show that IFN-β is highly effective in inhibiting virus replication in RPE cells. Specificity for TLR 3 signaling was demonstrated by the inhibition of poly I:C induction of IFN-β by pretreatment of RPE cells with anti-TLR 3 antibody.
Mucosal cells such as gastrointestinal, airway, and urinary epithelial cells are considered as the front line of defense against pathogenic microorganisms Schulz et al., 2002, Hornef et al., 2002, Pitman and Blumberg, 2000, Tsuboi et al., 2002 These cells have developed specific mechanisms for microbial protection that contribute to the innate immune response. Recently, it has been demonstrated that these epithelial cells contain several TLRs and respond to microbes by secreting cytokines and chemokines. For example, a murine intestinal epithelial cell line is highly responsive to LPS and expresses both TLR 4 and CD14. Corneal epithelial cells have been reported to express TLR 4 and CD14, and the LPS treatment of these cells resulted in the secretion of multiple proinflammatory cytokines and chemokines (Song et al., 2001). Like other epithelial cells, the RPE cell can also be considered as a front line defense against invading organisms. It is strategically located between the neural retina and the blood-rich choroid. The RPE forms a barrier, limiting access to photoreceptors and other neuronal cells within the retina. Earlier studies by our laboratory and others have revealed that RPE cells can be stimulated to produce a variety of cytokines, chemokines, and adhesion molecules Elner et al., 1990, Elner et al., 1992, Momma et al., 2003, Nagineni et al., 1994. In this report, we show that RPE cells possess a variety of TLRs and the costimulatory molecules, CD14 and MD2 (Tabeta et al., 2000). TLR 2 and TLR 4 are the most widely studied members of the TLR family Johnson et al., 2002, Faure et al., 2000, Opitz et al., 2001. Both of these TLRs were found on the RPE cell. Thus, RPE cells may defend against infections by sensing microbial invasion through multiple TLRs and the costimulatory molecules, CD14 and MD2.
To date, TLR 3 expression has been limited to a small number of cells types. Earlier studies demonstrate that TLR 3 is constitutively expressed on intestinal epithelial cells, dendritic cells, and mast cells Cario and Podolsky, 2000, Muzio et al., 2000. The dendritic cell is a potent antigen-presenting cell that expresses multiple TLRs including TLR 3 Banchereau et al., 2003, Jarrossay et al., 2001. Several studies suggest that TLR signaling on dendritic cells amplifies antigen presentation by producing proinflammatory cytokines, up-regulating co-stimulatory molecules, such as CD40, CD80, and CD86, and by increasing migration of cells to the lymph node (Palucka and Banchereau, 2002). The dendritic cell and the RPE cell share some common features. Both cells express TLR 3 and are APC. The dendritic cell responds to TLR 3 signaling by producing both IFN-α and IFN-β, whereas the RPE cell produces only IFN-β. This difference is probably a reflection of different cellular origins. Presently, dendritic cells are considered to be of bone marrow origin while RPE cells are of neural ectoderm origin. Additional studies are required to determine if the TLRs on the RPE cell also can augment APC functions.
The binding of dsRNA or poly I:C, an analog of dsRNA, to TLR 3 results in the production of type-1 IFNs and other cytokines and chemokines. Specificity for dsRNA interactions with TLR 3 was demonstrated by Matsumoto et al. (2002). Their studies pointed out that poly I:C-induced IFN-β was suppressed by pretreatment with monoclonal antibody to TLR 3. Additional studies on TLR 3 knockout mice revealed that poly I:C treatment up-regulated the production of type-1 IFNs (IFN-α, IFN-β) in wild-type mice but not in TLR 3 KO mice (Alexopoulou et al., 2001). In this report, we demonstrate that the poly I:C treatment of human RPE cells results in the production and release of IFN-β and not IFN-α. Moreover, these RPE cells are highly sensitive to the antiviral actions of human IFN-β. Clearly, dsRNA-mediated signaling in RPE cells can have a protective role in viral infections in the retina. In light of this, numerous in vivo and in vitro studies have identified that RPE cells are one of the principle targets for infectious agents, such as CMV, T. gondii, and murine coronaviruses Bodaghi et al., 1999, Detrick et al., 2001, Hooks et al., 1993, Nagineni et al., 1996. It is important to point out that additional TLRs may also participate in selected virus infections and further work is needed to better define this interaction Bieback et al., 2002, Compton et al., 2003, Kurt-Jones et al., 2000.
When an infecting agent enters the retina, it is critically important for the host to have a rapid response system to limit damage to nonregenerating retinal cells. The RPE cells are strategically placed to function as protective cells. Clearly, the innate immune system composed of TLRs is a primary rapid response system to infection. This study demonstrates that the RPE cell expresses TLRs 1–7, 9 and 10 and therefore can initiate signaling pathways that stimulate host defenses. Moreover, these cells respond to TLR stimulation by producing IFN-β, IL-6, IL-8, MCP-1, and sICAM-1. These cytokines, chemokines, and adhesion molecules together then participate in initiating adaptive immune responses. The demonstration that RPE cells express TLR 3 and release IFN-β represents a hitherto unrecognized biological role for the RPE cell.
References
- Alexopoulou L, Holt A.C, Medzhitov R, Flavell R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature. 2001;413:732–738. doi: 10.1038/35099560. [DOI] [PubMed] [Google Scholar]
- Banchereau J, Paczesny S, Blanco P, Bennett L, Pascual V, Fay J, Palucka A.K. Dendritic cells: controllers of the immune system and a new promise for immunotherapy. Ann. N.Y. Acad. Sci. 2003;987:180–187. doi: 10.1111/j.1749-6632.2003.tb06047.x. [DOI] [PubMed] [Google Scholar]
- Bauer S, Kirschning C.J, Hacker H, Redecke V, Hausmann S, Akira S, Wagner H, Lipford G.B. Human TLR9 confers responsiveness to bacterial DNA via species-specific CpG motif recognition. Proc. Natl. Acad. Sci. U. S. A. 2001;98:9237–9242. doi: 10.1073/pnas.161293498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bieback K, Lien E, Klagge I.M, Avota E, Schneider-Schaulies J, Duprex W.P, Wagner H, Kirschning C.J, Ter Meulen V, Schneider-Schaulies S. Hemagglutinin protein of wild-type measles virus activates toll-like receptor 2 signaling. J. Virol. 2002;76:8729–8736. doi: 10.1128/JVI.76.17.8729-8736.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bodaghi B, Slobbe-van Drunen M.E, Topilko A, Perret E, Vossen R.C, van Dam-Mieras M.C, Zipeto D, Virelizier J.L, LeHoang P, Bruggeman C.A, Michelson S. Entry of human cytomegalovirus into retinal pigment epithelial and endothelial cells by endocytosis. Investig. Ophthalmol. Vis. Sci. 1999;40:2598–2607. [PubMed] [Google Scholar]
- Bok D. Retinal photoreceptor-pigment epithelium interactions. Friedenwald lecture. Investig. Ophthalmol. Vis. Sci. 1985;26:1659–1694. [PubMed] [Google Scholar]
- Bok D. The retinal pigment epithelium: a versatile partner in vision. J. Cell Sci., Suppl., Suppl. 1993;17:189–195. doi: 10.1242/jcs.1993.supplement_17.27. [DOI] [PubMed] [Google Scholar]
- Cai J, Nelson K.C, Wu M, Sternberg P, Jr., Jones D.P. Oxidative damage and protection of the RPE. Prog. Retin. Eye Res. 2000;19:205–221. doi: 10.1016/s1350-9462(99)00009-9. [DOI] [PubMed] [Google Scholar]
- Campos M.A, Almeida I.C, Takeuchi O, Akira S, Valente E.P, Procopio D.O, Travassos L.R, Smith J.A, Golenbock D.T, Gazzinelli R.T. Activation of Toll-like receptor-2 by glycosylphosphatidylinositol anchors from a protozoan parasite. J. Immunol. 2001;167:416–423. doi: 10.4049/jimmunol.167.1.416. [DOI] [PubMed] [Google Scholar]
- Cario E, Podolsky D.K. Differential alteration in intestinal epithelial cell expression of toll-like receptor 3 (TLR3) and TLR4 in inflammatory bowel disease. Infect. Immun. 2000;68:7010–7017. doi: 10.1128/iai.68.12.7010-7017.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chin M.S, Nagineni C.N, Hooper L.C, Detrick B, Hooks J.J. Cyclooxygenase-2 gene expression and regulation in human retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2001;42:2338–2346. [PubMed] [Google Scholar]
- Compton T, Kurt-Jones E.A, Boehme K.W, Belko J, Latz E, Golenbock D.T, Finberg R.W. Human cytomegalovirus activates inflammatory cytokine responses via CD14 and Toll-like receptor 2. J. Virol. 2003;77:4588–4596. doi: 10.1128/JVI.77.8.4588-4596.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Detrick B, Rhame J, Wang Y, Nagineni C.N, Hooks J.J. Cytomegalovirus replication in human retinal pigment epithelial cells. Altered expression of viral early proteins. Investig. Ophthalmol. Vis. Sci. 1996;37:814–825. [PubMed] [Google Scholar]
- Detrick B, Nagineni C.N, Grillone L.R, Anderson K.P, Henry S.P, Hooks J.J. Inhibition of human cytomegalovirus replication in a human retinal epithelial cell model by antisense oligonucleotides. Investig. Ophthalmol. Vis. Sci. 2001;42:163–169. [PubMed] [Google Scholar]
- Elner V.M, Strieter R.M, Elner S.G, Baggiolini M, Lindley I, Kunkel S.L. Neutrophil chemotactic factor (IL-8) gene expression by cytokine-treated retinal pigment epithelial cells. Am. J. Pathol. 1990;136:745–750. [PMC free article] [PubMed] [Google Scholar]
- Elner V.M, Scales W, Elner S.G, Danforth J, Kunkel S.L, Strieter R.M. Interleukin-6 (IL-6) gene expression and secretion by cytokine-stimulated human retinal pigment epithelial cells. Exp. Eye Res. 1992;54:361–368. doi: 10.1016/0014-4835(92)90048-w. [DOI] [PubMed] [Google Scholar]
- Faure E, Equils O, Sieling P.A, Thomas L, Zhang F.X, Kirschning C.J, Polentarutti N, Muzio M, Arditi M. Bacterial lipopolysaccharide activates NF-kappaB through toll-like receptor 4 (TLR-4) in cultured human dermal endothelial cells. Differential expression of TLR-4 and TLR-2 in endothelial cells. J. Biol. Chem. 2000;275:11058–11063. doi: 10.1074/jbc.275.15.11058. [DOI] [PubMed] [Google Scholar]
- Gewirtz A.T, Navas T.A, Lyons S, Godowski P.J, Madara J.L. Cutting edge: bacterial flagellin activates basolaterally expressed TLR5 to induce epithelial proinflammatory gene expression. J. Immunol. 2001;167:1882–1885. doi: 10.4049/jimmunol.167.4.1882. [DOI] [PubMed] [Google Scholar]
- Gordon S. Pattern recognition receptors: doubling up for the innate immune response. Cell. 2002;111:927–930. doi: 10.1016/s0092-8674(02)01201-1. [DOI] [PubMed] [Google Scholar]
- Hamel C.P, Tsilou E, Harris E, Pfeffer B.A, Hooks J.J, Detrick B, Redmond T.M. A developmentally regulated microsomal protein specific for the pigment epithelium of the vertebrate retina. J. Neurosci. Res. 1993;34:414–425. doi: 10.1002/jnr.490340406. [DOI] [PubMed] [Google Scholar]
- Hooks J.J, Percopo C, Wang Y, Detrick B. Retina and retinal pigment epithelial cell autoantibodies are produced during murine coronavirus retinopathy. J. Immunol. 1993;151:3381–3389. [PubMed] [Google Scholar]
- Hornef M.W, Frisan T, Vandewalle A, Normark S, Richter-Dahlfors A. Toll-like receptor 4 resides in the Golgi apparatus and colocalizes with internalized lipopolysaccharide in intestinal epithelial cells. J. Exp. Med. 2002;195:559–570. doi: 10.1084/jem.20011788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito T, Amakawa R, Kaisho T, Hemmi H, Tajima K, Uehira K, Ozaki Y, Tomizawa H, Akira S, Fukuhara S. Interferon-alpha and interleukin-12 are induced differentially by Toll-like receptor 7 ligands in human blood dendritic cell subsets. J. Exp. Med. 2002;195:1507–1512. doi: 10.1084/jem.20020207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janssens S, Beyaert R. Role of Toll-like receptors in pathogen recognition. Clin. Microbiol. Rev. 2003;16:637–646. doi: 10.1128/CMR.16.4.637-646.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarrossay D, Napolitani G, Colonna M, Sallusto F, Lanzavecchia A. Specialization and complementarity in microbial molecule recognition by human myeloid and plasmacytoid dendritic cells. Eur. J. Immunol. 2001;31:3388–3393. doi: 10.1002/1521-4141(200111)31:11<3388::aid-immu3388>3.0.co;2-q. [DOI] [PubMed] [Google Scholar]
- Johnson G.B, Brunn G.J, Kodaira Y, Platt J.L. Receptor-mediated monitoring of tissue well-being via detection of soluble heparan sulfate by Toll-like receptor 4. J. Immunol. 2002;168:5233–5239. doi: 10.4049/jimmunol.168.10.5233. [DOI] [PubMed] [Google Scholar]
- Kurt-Jones E.A, Popova L, Kwinn L, Haynes L.M, Jones L.P, Tripp R.A, Walsh E.E, Freeman M.W, Golenbock D.T, Anderson L.J, Finberg R.W. Pattern recognition receptors TLR4 and CD14 mediate response to respiratory syncytial virus. Nat. Immunol. 2000;1:398–401. doi: 10.1038/80833. [DOI] [PubMed] [Google Scholar]
- Li W, Nagineni C.N, Hooks J.J, Chepelinsky A.B, Egwuagu C.E. Interferon-gamma signaling in human retinal pigment epithelial cells mediated by STAT1, ICSBP, and IRF-1 transcription factors. Investig. Ophthalmol. Vis. Sci. 1999;40:976–982. [PubMed] [Google Scholar]
- Lutty G, Grunwald J, Majji A.B, Uyama M, Yoneya S. Changes in choriocapillaris and retinal pigment epithelium in age-related macular degeneration. Mol. Vis. 1999;5:35. [PubMed] [Google Scholar]
- Matsumoto M, Kikkawa S, Kohase M, Miyake K, Seya T. Establishment of a monoclonal antibody against human Toll-like receptor 3 that blocks double-stranded RNA-mediated signaling. Biochem. Biophys. Res. Commun. 2002;293:1364–1369. doi: 10.1016/S0006-291X(02)00380-7. [DOI] [PubMed] [Google Scholar]
- Medzhitov R, Janeway C.A., Jr. Innate immunity: the virtues of a nonclonal system of recognition. Cell. 1997;91:295–298. doi: 10.1016/s0092-8674(00)80412-2. [DOI] [PubMed] [Google Scholar]
- Momma Y, Nagineni C.N, Chin M.S, Srinivasan K, Detrick B, Hooks J.J. Differential expression of chemokines by human retinal pigment epithelial cells infected with cytomegalovirus. Investig. Ophthalmol. Vis. Sci. 2003;44:2026–2033. doi: 10.1167/iovs.02-0980. [DOI] [PubMed] [Google Scholar]
- Morimura H, Fishman G.A, Grover S.A, Fulton A.B, Berson E.L, Dryja T.P. Mutations in the RPE65 gene in patients with autosomal recessive retinitis pigmentosa or leber congenital amaurosis. Proc. Natl. Acad. Sci. U. S. A. 1998;95:3088–3093. doi: 10.1073/pnas.95.6.3088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morimura H, Saindelle-Ribeaudeau F, Berson E.L, Dryja T.P. Mutations in RGR, encoding a light-sensitive opsin homologue, in patients with retinitis pigmentosa. Nat. Genet. 1999;23:393–394. doi: 10.1038/70496. [DOI] [PubMed] [Google Scholar]
- Muzio M, Bosisio D, Polentarutti N, D'Amico G, Stoppacciaro A, Mancinelli R, van't Veer C, Penton-Rol G, Ruco L.P, Allavena P, Mantovani A. Differential expression and regulation of toll-like receptors (TLR) in human leukocytes: selective expression of TLR3 in dendritic cells. J. Immunol. 2000;164:5998–6004. doi: 10.4049/jimmunol.164.11.5998. [DOI] [PubMed] [Google Scholar]
- Nagineni C.N, Detrick B, Hooks J.J. Synergistic effects of gamma interferon on inflammatory mediators that induce interleukin-6 gene expression and secretion by human retinal pigment epithelial cells. Clin. Diagn. Lab. Immunol. 1994;1:569–577. doi: 10.1128/cdli.1.5.569-577.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagineni C.N, Pardhasaradhi K, Martins M.C, Detrick B, Hooks J.J. Mechanisms of interferon-induced inhibition of Toxoplasma gondii replication in human retinal pigment epithelial cells. Infect. Immun. 1996;64:4188–4196. doi: 10.1128/iai.64.10.4188-4196.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagineni C.N, Detrick B, Hooks J.J. Toxoplasma gondii infection induces gene expression and secretion of interleukin 1 (IL-1), IL-6, granulocyte-macrophage colony-stimulating factor, and intercellular adhesion molecule 1 by human retinal pigment epithelial cells. Infect. Immun. 2000;68:407–410. doi: 10.1128/iai.68.1.407-410.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagineni C.N, Detrick B, Hooks J.J. Transforming growth factor-beta expression in human retinal pigment epithelial cells is enhanced by Toxoplasma gondii: a possible role in the immunopathogenesis of retinochoroiditis. Clin. Exp. Immunol. 2002;128:372–378. doi: 10.1046/j.1365-2249.2002.01815.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Opitz B, Schroder N.W, Spreitzer I, Michelsen K.S, Kirschning C.J, Hallatschek W, Zahringer U, Hartung T, Gobel U.B, Schumann R.R. Toll-like receptor-2 mediates Treponema glycolipid and lipoteichoic acid-induced NF-kappaB translocation. J. Biol. Chem. 2001;276:22041–22047. doi: 10.1074/jbc.M010481200. [DOI] [PubMed] [Google Scholar]
- Palucka K, Banchereau J. How dendritic cells and microbes interact to elicit or subvert protective immune responses. Curr. Opin. Immunol. 2002;14:420–431. doi: 10.1016/s0952-7915(02)00365-5. [DOI] [PubMed] [Google Scholar]
- Percopo C.M, Hooks J.J, Shinohara T, Caspi R, Detrick B. Cytokine-mediated activation of a neuronal retinal resident cell provokes antigen presentation. J. Immunol. 1990;145:4101–4107. [PubMed] [Google Scholar]
- Pitman R.S, Blumberg R.S. First line of defense: the role of the intestinal epithelium as an active component of the mucosal immune system. J. Gastroenterol. 2000;35:805–814. doi: 10.1007/s005350070017. [DOI] [PubMed] [Google Scholar]
- Schjetne K.W, Thompson K.M, Nilsen N, Flo T.H, Fleckenstein B, Iversen J.G, Espevik T, Bogen B. Cutting Edge: link between innate and adaptive immunity: Toll-like receptor 2 internalizes antigen for presentation to CD4+T cells and could be an efficient vaccine target. J. Immunol. 2003;171:32–36. doi: 10.4049/jimmunol.171.1.32. [DOI] [PubMed] [Google Scholar]
- Schnare M, Barton G.M, Holt A.C, Takeda K, Akira S, Medzhitov R. Toll-like receptors control activation of adaptive immune responses. Nat. Immunol. 2001;2:947–950. doi: 10.1038/ni712. [DOI] [PubMed] [Google Scholar]
- Schulz C, Farkas L, Wolf K, Kratzel K, Eissner G, Pfeifer M. Differences in LPS-induced activation of bronchial epithelial cells (BEAS-2B) and type II-like pneumocytes (A-549) Scand. J. Immunol. 2002;56:294–302. doi: 10.1046/j.1365-3083.2002.01137.x. [DOI] [PubMed] [Google Scholar]
- Song P.I, Abraham T.A, Park Y, Zivony A.S, Harten B, Edelhauser H.F, Ward S.L, Armstrong C.A, Ansel J.C. The expression of functional LPS receptor proteins CD14 and toll-like receptor 4 in human corneal cells. Investig. Ophthalmol. Vis. Sci. 2001;42:2867–2877. [PubMed] [Google Scholar]
- Tabeta K, Yamazaki K, Akashi S, Miyake K, Kumada H, Umemoto T, Yoshie H. Toll-like receptors confer responsiveness to lipopolysaccharide from Porphyromonas gingivalis in human gingival fibroblasts. Infect. Immun. 2000;68:3731–3735. doi: 10.1128/iai.68.6.3731-3735.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takeda K, Kaisho T, Akira S. Toll-like receptors. Annu. Rev. Immunol. 2003;21:335–376. doi: 10.1146/annurev.immunol.21.120601.141126. [DOI] [PubMed] [Google Scholar]
- Tsuboi N, Yoshikai Y, Matsuo S, Kikuchi T, Iwami K, Nagai Y, Takeuchi O, Akira S, Matsuguchi T. Roles of toll-like receptors in C-C chemokine production by renal tubular epithelial cells. J. Immunol. 2002;169:2026–2033. doi: 10.4049/jimmunol.169.4.2026. [DOI] [PubMed] [Google Scholar]
- Zuany-Amorim C, Hastewell J, Walker C. Toll-like receptors as potential therapeutic targets for multiple diseases. Nat. Rev. Drug Discov. 2002;1:797–807. doi: 10.1038/nrd914. [DOI] [PubMed] [Google Scholar]