

Abstract
Angiogenesis, a process of new blood vessel formation, is a prerequisite for tumour growth to supply the proliferating tumour with oxygen and nutrients. The angiogenic process may contribute to tumour progression, invasion and metastasis, and is generally accepted as an indicator of tumour prognosis. Therefore, targeting tumour angiogenesis has become of high clinical relevance. The current review aimed to highlight mechanistic details of anti-angiogenic therapies and how they relate to classification and treatment rationales. Angiogenesis inhibitors are classified into either direct inhibitors that target endothelial cells in the growing vasculature or indirect inhibitors that prevent the expression or block the activity of angiogenesis inducers. The latter class extends to include targeted therapy against oncogenes, conventional chemotherapeutic agents and drugs targeting other cells of the tumour micro-environment. Angiogenesis inhibitors may be used as either monotherapy or in combination with other anticancer drugs. In this context, many preclinical and clinical studies revealed higher therapeutic effectiveness of the combined treatments compared with individual treatments. The proper understanding of synergistic treatment modalities of angiogenesis inhibitors as well as their wide range of cellular targets could provide effective tools for future therapies of many types of cancer.
Keywords: angiogenesis inhibitors, cancer therapy, tumour micro-environment
Introduction
Two major processes of blood vessel formation are implicated in the development of vascular system: vasculogenesis and angiogenesis. Vasculogenesis prevails in the embryo and refers to the formation of de novo blood vessels by in situ differentiation of the mesoderm-derived angioblasts and endothelial precursors. Angiogenesis is the formation of new capillaries from pre-existing vessels and circulating endothelial precursors (Polverini, 2002; Chung et al., 2010; Ribatti and Djonov, 2012). Angiogenesis is a tightly controlled dynamic process that can occur physiologically in those tissues that undergo active remodelling in response to stress and hypoxia (Carmeliet, 2003; Folkman, 2007). However, it can be aberrantly activated during many pathological conditions such as cancer, diabetic retinopathy as well as numerous ischaemic, inflammatory, infectious and immune disorders (Carmeliet, 2003; Ali and El-Remessy, 2009; Willis et al., 2011). Although the concept of proposing angiogenesis inhibitors as anticancer drugs received considerable skepticism when first presented by Dr. Folkman in the early 1970s (Folkman, 1971), active research in the field and subsequent clinical trials eventually resulted in US Food and Drug Administration (FDA) approval of bevacizumab for colorectal cancer in 2004 (Cohen et al., 2007). Since then, several angiogenic inhibitors have been identified. This review will provide an overview of the key mechanisms involved in tumour angiogenesis, classification of angiogenesis inhibitors as well as treatment rationales from the mechanistic point of view.
Sustained angiogenesis as a hallmark of cancer
Proliferating tumours tend to activate an angiogenic phenotype to fulfil their increased demand of oxygen and nutrients (Hanahan and Folkman, 1996; Carmeliet, 2005). Additionally, paracrine release of anti-apoptotic factors from activated endothelial cells in the newly formed vasculature supplies tumour cells with a survival privilege (Folkman, 2003). Consequently, in order to progress, tumours tend to activate an event called ‘angiogenic switch’ by shifting the balance of endogenous angiogenesis inducers and inhibitors towards a pro-angiogenic outcome. As a result, dormant lesion progresses into outgrowing vascularized tumour and eventually into a malignant phenotype (Hanahan and Folkman, 1996; Baeriswyl and Christofori, 2009). Hypoxia drives such imbalance through up-regulation of the transcription factor hypoxia inducible factor-1α (HIF-1α), which in turn increases the expression of many angiogenesis inducers as well as suppresses the expression of endogenous angiogenesis inhibitors (Pugh and Ratcliffe, 2003). In spite of that, accumulating evidence indicates that angiogenic cascade can be also driven by alternative HIF-1-independent pathways (Mizukami et al., 2007; Arany et al., 2008; Lee, 2013).
As summarized in Table 1, the angiogenesis inducers are a wide range of mediators that include many growth factors, a plethora of cytokines, bioactive lipids, matrix-degrading enzymes and a number of small molecules (Folkman, 1995; Folkman, 2003; Lopez-Lopez et al., 2004; Bouis et al., 2006; El-Remessy et al., 2007; Bid et al., 2011; MacLauchlan et al., 2011; Murakami, 2011; Fagiani and Christofori, 2013; Qin et al., 2013). Pro-angiogenic growth factors mostly activate a series of surface receptors in a series of paracrine and autocrine loops with the VEGF-A signalling representing the critical rate-limiting step, physiologically and pathologically. VEGF-A (traditionally known as VEGF) is the most potent VEGF isoform that acts mainly on VEGF receptor 2 (VEGFR2) to mediate vascular permeability, endothelial proliferation, migration and survival (Takahashi and Shibuya, 2005; Bouis et al., 2006). In spite of the well-established master roles of VEGF signalling in literature, those processes are probably accomplished through a highly regulated interplay between VEGF and other pro-angiogenic factors. In this context, basic fibroblast growth factor (bFGF) activation of the endothelium is required for maintenance of VEGFR2 expression and the ability to respond to VEGF stimulation (Murakami et al., 2011). Similarly, sphingosine-1-phosphate (S1P), a pleiotropic bioactive lipid that can directly contribute to tumour angiogenesis (reviewed in Sabbadini, 2011), is needed for VEGF-induced blood vessel formation, indicating the cooperation between S1P and VEGF in tumour angiogenesis (Visentin et al., 2006). As a net result, the pro-angiogenic interplay of those ligands and others dominates over the activities of two dozen endogenous angiogenesis inhibitors that can be either matrix-derived inhibitors or non–matrix-derived inhibitors (Nyberg et al., 2005).
Table 1.
Pro-angiogenic mediators implicated in tumour angiogenesis
| Category | Examples | References |
|---|---|---|
| Growth factors | VEGFs | Bouis et al., 2006 |
| FGFs | Bouis et al., 2006 | |
| TGFs | Bouis et al., 2006 | |
| PDGFs | Bouis et al., 2006 | |
| Insulin-like growth factors | Lopez-Lopez et al., 2004; Bid et al., 2011 | |
| ANGs | Fagiani and Christofori, 2013 | |
| Cytokines | IL-8 | Strieter et al., 2004 |
| CSF-1 | Lin et al., 2006 | |
| Bioactive lipids | PGE2 | Wang and Dubois, 2010 |
| S1P | Murakami, 2011 | |
| Matrix-degrading enzymes | MMPs | Bourboulia and Stetler-Stevenson, 2010 |
| Heparanases | Vlodavsky and Friedmann, 2001 | |
| Small mediators | NO | MacLauchlan et al., 2011 |
| Peroxynitrite | El-Remessy et al., 2007 | |
| Serotonin | Qin et al., 2013 | |
| Histamine | Qin et al., 2013 |
The multistep angiogenic process starts with vasodilation and increased permeability of existing vessels in response to tumour cell-secreted VEGF. This is accompanied by loosening of pericytes covering mediated by angiopoietin-2 (ANG2), a ligand of tyrosine kinase with immunoglobulin-like and EGF-like domains 2 (TIE2) receptor (Bergers and Benjamin, 2003; Jain, 2003; Fagiani and Christofori, 2013). Meanwhile, many secreted matrix-degrading enzymes, such as MMPs and heparanases, function in concert to dissolve the basement membrane and to remodel the extracellular matrix (ECM) as well as to liberate more pro-angiogenic growth factors (bFGF and VEGF) from matrix heparan sulfate proteoglycans (HSPGs) respectively (Houck et al., 1992; Whitelock et al., 1996; Vlodavsky and Friedmann, 2001; Tang et al., 2005; van Hinsbergh and Koolwijk, 2008). The overall chemotactic angiogenic stimuli guide endothelial cells to migrate, to align into tube-like structures and to eventually form new blood vessels. However, such blood vessels are characterized by being disorganized, chaotic, haemorrhagic and poorly functioning (Bergers and Benjamin, 2003).
The angiogenic phenotype in tumour micro-environment can further be sustained and extravagated by the recruitment of other types of stromal cells. Stromal cells such as fibroblasts, mesenchymal stem cells and various bone marrow-derived myeloid cells including macrophages, TIE2-expressing monocytes, neutrophils and mast cells contribute to tumour angiogenesis through their production of growth factors, cytokines and proteases (Murdoch et al., 2008; Joyce and Pollard, 2009; Cirri and Chiarugi, 2011). For example, in response to cancer cell-derived TGF-β, PDGF or bFGF, fibroblasts are transformed to an activated phenotype with a higher proliferative activity and myofibroblastic characteristics (Kalluri and Zeisberg, 2006; Cirri and Chiarugi, 2011). Such carcinoma-associated fibroblasts (CAFs) were shown to promote angiogenesis and metastasis by secreting large amounts of MMP-2 and MMP-9 as well as by expressing many cytokines and chemokines that resulted in immune cell infiltration (Gerber et al., 2009; Giannoni et al., 2010). Furthermore, it has been shown that PDGF-C produced by CAFs is able to elicit VEGF production from tumour cells, thereby sustaining the angiogenic shift (Crawford et al., 2009). Similarly, tumour-associated macrophages (TAMs), one of the bone marrow myeloid-derived cells, are induced to develop into polarized type II (alternatively activated or M2 macrophages), upon exposure to tumour hypoxia and tumour cell-derived cytokines (Leek et al., 2002; Rogers and Holen, 2011). M2 macrophages tend to produce many pro-angiogenic growth factors, cytokines and matrix-degrading enzymes such as VEGF, PDGF, bFGF, TNF-α, COX-2, MMP-9, MMP-7 and MMP-12 (Lewis and Pollard, 2006).
From another perspective, angiogenesis may be dispensable for progression of some malignancies. For example, some tumours may co-opt pre-existent vessels as an alternative way to obtain blood supply. Vessel co-option was first described in the brain, one of the most densely vascularized organs, in which tumours may develop in earlier stages without the activation of angiogenic response (Holash et al., 1999; Leenders et al., 2002; Bergers and Benjamin, 2003; Hillen and Griffioen, 2007). In another example, hypovascularized tumours such as pancreatic ductal adenocarcinoma may involve certain adaptation to flourish in the absence of prominent angiogenesis (Bergers and Hanahan, 2008). Obviously, in both cases, tumours may be intrinsically indifferent to angiogenesis inhibitors. However, in most other cases, therapy directed towards the vasculature of solid tumours is being considered as an important direction in cancer treatment.
Classification of angiogenesis inhibitors
Growth of newly formed vessels in tumour micro-environment can be inhibited directly by targeting endothelial cells in the growing vasculature or indirectly by targeting either tumour cells or the other tumour-associated stromal cells. Therefore, angiogenesis inhibitors can be classified into direct and indirect inhibitors (Kerbel and Folkman, 2002; Folkman, 2007).
Direct endogenous inhibitors of angiogenesis
Direct endogenous inhibitors of angiogenesis, such as angiostatin, endostatin, arrestin, canstatin, tumstatin and others, are fragments released on proteolysis of distinct ECM molecules. Endogenous inhibitors prevent vascular endothelial cells from proliferating, migrating in response to a spectrum of angiogenesis inducers, including VEGF, bFGF, IL-8 and PDGF (Kerbel and Folkman, 2002; Abdollahi et al., 2004; Mundel and Kalluri, 2007; Ribatti, 2009). This direct anti-angiogenic effect may be mediated by interference with endothelial integrins along with several intracellular signalling pathways (Mundel and Kalluri, 2007). For example, the ability of tumstatin-derived active peptide to inhibit angiogenesis and tumour growth is associated with the expression of the adhesion receptor, αvβ3 integrin, on tumour endothelial cells (Eikesdal et al., 2008). Through binding αvβ3 integrin, full tumstatin was found to inhibit endothelial cell activation of focal adhesion kinase, PI3K, Akt, mammalian target of rapamycin (mTOR) and others (Maeshima et al., 2002). Direct targeting of those signalling pathways by endogenous inhibitors was thought to be the least likely to induce acquired drug resistance because they target endothelial cells with assumed genetic stability rather than unstable mutating tumour cells (Kerbel and Folkman, 2002). However, endostatin has not yet led to any documented benefit to patients in randomized phase III trials, or even modest activity in phase II trials (Ellis and Hicklin, 2008).
Indirect inhibitors of angiogenesis
Indirect inhibitors of angiogenesis classically prevent the expression or block the activity of pro-angiogenic proteins (Folkman, 2007). For example, Iressa, an EGF receptor (EGFR) TK inhibitor (TKI), blocks tumour expression of many pro-angiogenic factors; bevacizumab, a monoclonal antibody, neutralizes VEGF after its secretion from tumour cells whereas sunitinib, a multiple receptor TKI, blocks the endothelial cell receptors (VEGFR1, VEGFR2 and VEGFR3), preventing their response to the secreted VEGF (Folkman, 2007; Roskoski, 2007). In addition, this class extends to include conventional chemotherapeutic agents, targeted therapy against oncogenes and drugs targeting other cells of the tumour micro-environment (Kerbel et al., 2000; Ferrara and Kerbel, 2005).
Conventional chemotherapeutic agents
Conventional chemotherapeutic agents have been shown to have anti-angiogenic properties in addition to the ability to induce direct cancer cell death. Such chemotherapeutic agents can affect the endothelial cell population in the tumour bed during treatment cycles because they have significantly higher proliferation rates than resting endothelium outside a tumour, making them more susceptible to cytotoxic effect (Kerbel et al., 2000; Folkman, 2003). However, the cyclic treatment rationale of cytotoxic drugs allows the potential damage to the tumour vasculature to be repaired during the long breaks. Thus, continuous low doses of chemotherapeutic agents were suggested as a way to reduce side effects and drug resistance (Drevs et al., 2004). This modality is termed metronomic therapy, and clinically, it refers to the daily administration of 5–10% of the phase II-recommended dose of the chemotherapeutic agent (Penel et al., 2012). The extended use of such low doses of cytotoxic agents elicits an anti-angiogenic activity through induction of endothelial cell apoptosis and decreasing the level of circulating endothelial precursors (Hamano et al., 2004; Shahrzad et al., 2008). In clinical investigations, metronomic dosing of cyclophosphamide and others showed promising efficacy in patients with advanced, multiple metastasized and/or multiple pretreated solid tumours (Lord et al., 2007; Fontana et al., 2010; Nelius et al., 2011; Gebbia et al., 2012; Briasoulis et al., 2013; Navid et al., 2013).
VEGF-targeted therapy
VEGF-targeted therapy includes neutralizing antibodies to VEGF (e.g. bevacizumab) or VEGFRs (e.g. ramucirumab), soluble VEGFR/VEGFR hybrids (e.g. VEGF-Trap) and TKIs with selectivity for VEGFRs (e.g. sunitinib and sorafenib; Baka et al., 2006; Ellis and Hicklin, 2008; Hsu and Wakelee, 2009). Bevacizumab, a humanized monoclonal antibody against all isoforms of VEGF-A, has been approved for the treatment of colorectal, lung, glioblastoma and renal cell carcinoma (Hsu and Wakelee, 2009). Many other clinical trials with promising efficacy were also conducted in other cancers such as head and neck cancer, hepatocellular carcinoma, ovarian cancer, metastatic melanoma and gastric cancer (Argiris et al., 2011; 2013; Burger et al., 2011; Ohtsu et al., 2011; Fang et al., 2012; Minor, 2012; Schuster et al., 2012; Van Cutsem et al., 2012). However, for metastatic breast cancer, bevacizumab had been initially granted an accelerated FDA approval, which was later withdrawn due to lack of improvement evidence in disease-related symptoms or overall survival (Burstein, 2011; Montero et al., 2012). Similarly, clinical trials showed that the addition of bevacizumab to the treatment regimens of advanced pancreatic cancer did not extend overall survival (Chiu and Yau, 2012). The neutralization of VEGF-A can also be achieved by soluble receptor construct (VEGF-Trap) that monomerically ‘traps’ the different isoforms of VEGF-A, in addition to VEGF-B and placental growth factor (Rudge et al., 2007). VEGF-Trap showed clinical benefit in a phase III trial of oxaliplatin pretreated metastatic patients with colorectal cancer, and is currently being investigated in a prostate cancer phase III trial (Gaya and Tse, 2012). TKIs are small molecules with different chemical structures that have the ability to interact physically with the highly conserved kinase domain shared by different VEGFRs as well as PDGF receptors (PDGFRs), FGF receptors (FGFRs), EGFR, Raf kinases and c-Kit (a receptor of the pluripotent cell growth factor, stem cell factor). Such interaction directly inhibits tyrosine phosphorylation and the subsequent many downstream pro-angiogenic signalling networks (Baka et al., 2006; Ivy et al., 2009). Those multi-targeted TKIs demonstrated efficacy against various solid malignancies in different clinical trials, some of which have lead eventually to FDA approval of sunitinib and sorafenib. Sunitinib, known to inhibit several receptor TKs (RTKs) including VEGFR1–3, PDGFR-α, PDGFR-β, c-Kit, colony-stimulating factor-1 receptor (CSF-1R) and Flt-3, was approved for the treatment of renal cell carcinoma and gastrointestinal stromal cell tumours. Sorafenib that acts also by inhibiting VEGFR1–3 and PDGFR-β in addition to the serine–threonine kinases Raf-1, B-Raf, was approved for hepatocellular carcinoma in addition to renal cell carcinoma (Llovet et al., 2008; Ivy et al., 2009; Huang et al., 2010).
FGF-targeted therapies
FGF-targeted therapies were recently reconsidered as promising anti-angiogenic and anti-tumour agents after a long period of little attention for drug development, partly due to redundancy (Bono et al., 2013). The FGFR superfamily with its 18 ligands and four receptors has been involved in endothelial cell migration, proliferation and differentiation (Presta et al., 2005). Therapeutic targeting of FGF/FGFR signalling was accomplished by either monoclonal antibodies that inhibit FGFs binding, small molecules that inhibit FGFR TK activity or allosteric modulators that bind the extracellular FGFR domain. Monoclonal antibodies against bFGF displayed potent anti-tumour and anti-angiogenic effects in different preclinical cancer models, which warrant further clinical evaluation (Zhao et al., 2010; Wang et al., 2012). Pan inhibitors of the FGFR TKs such as AZD4547 (blocks the activity of FGFR1–3) and ponatinib (blocks all the FGFR isoforms) elicited potent anti-tumour activities in preclinical investigations so they are currently being evaluated in clinical trials. Those inhibitors displayed the greatest potency in FGFR-driven cancer models, which may be attributed to the interference with the oncogenic functions of either amplified or constitutively active FGFR (Dutt et al., 2011; Zhao et al., 2011; Gavine et al., 2012; Gozgit et al., 2012). Accordingly, further studies are needed to evaluate the relative contribution of angiogenic versus oncogenic inhibitory mechanisms towards the overall anti-tumour activity. The allosteric antagonist of the FGFR, SSR128129E, showed a strong anti-angiogenic activity in addition to tumour growth and metastasis inhibitory effects in animal models of arthritis and cancer respectively. Because allosteric modulators leave a residual level of baseline signalling, they have the ability to fine-tune target biological responses. As a result, allosteric multi-FGFR inhibitors may have an improved benefit/risk ratio that is not attainable with the other TKIs (Bono et al., 2013; Herbert et al., 2013). However, preclinical findings suggest that long-term clinical outcomes may improve with blockade of additional pro-angiogenic RTKs that may also reduce the risk of drug resistance (Hilberg et al., 2008). For example, dual inhibition of VEGFRs and FGFRs using brivanib produced enduring tumour stasis and angiogenic blockade following the failure of VEGF-targeted therapies (Allen et al., 2011). Furthermore, triple inhibition of FGFRs, VEGFRs and PDGFR(s) using dovitinib (TKI258) or nintedanib (BIBF 1120) displayed broad-spectrum anti-tumour activities in several tumour xenograft models as well as promising data in clinical trials. Combined inhibition of FGFR/VEGFR/PDGFR targets not only tumour cells, but also endothelial cells, pericytes and smooth muscle cells, resulting in an effective inhibition of tumour growth, angiogenesis and metastasis even in advanced tumour stages (Hilberg et al., 2008; Ledermann et al., 2011; Taeger et al., 2011; Chen et al., 2012; Angevin et al., 2013).
Oncogene-targeted therapy
Oncogenes, genes that cause the transformation of normal cells into cancerous cells, are thought to up-regulate many pro-angiogenic proteins. Therefore, anticancer drugs that were developed for their capacity to block an oncogene also have an indirect anti-angiogenic activity (Kerbel et al., 2000; Bergers and Benjamin, 2003; Folkman, 2003). For example, dasatinib and other inhibitors of sarcoma (Src), an aberrantly activated non-RTK associated with many human malignancies, showed potent anti-angiogenic effects through the down-regulation of VEGF and IL-8 (Summy et al., 2005; Han et al., 2006; Haura et al., 2010). Another example is to target the oncogenic Ras using farnesyl transferase (FT) inhibitors, which inhibit post-translational farnesylation of Ras that governs the latter's activity (Awada et al., 2002). FT inhibitors were found to inhibit tumour VEGF expression and block FTase-dependent Ras activation, which is critically involved in VEGF-elicited angiogenic signal transduction and angiogenesis (Han et al., 2005; Izbicka et al., 2005; Kim et al., 2010). In addition to classical oncogenes inhibition, interference with other tumour-deregulated signalling pathways would offer another approach in targeting angiogenesis. For example, inhibitors of heat shock protein 90 (HSP90), a chaperone molecule known to protect oncoproteins from misfolding and degradation in the protein-rich intracellular environment, were found to prevent VEGF production and to disrupt multiple pro-angiogenic signalling pathways in numerous cancer cells. They were also shown to inhibit tumour growth and vascularity of different human tumour xenografts (Sanderson et al., 2006; Lang et al., 2007; Eccles et al., 2008; Trepel et al., 2010; Moser et al., 2012). Proteasome inhibitors, such as bortezomib (PS-341) or MG-132, were also shown to reduce tumour growth and vascularity of squamous cell carcinoma and pancreatic cancer xenograft probably through inhibition of NF–κB-dependent release of pro-angiogenic gene products, VEGF and IL-8 (Sunwoo et al., 2001; Nawrocki et al., 2002; Matsuo et al., 2009). Similarly, inhibition of B-cell lymphoma 2 (Bcl-2), a prosurvival protein that regulates apoptosis by preventing the mitochondrial release of pro-apoptogenic factors, was shown to prevent NF-κB-mediated release of the pro-angiogenic factors IL-8 and CXC chemokine ligand 1 (CXCL1) as well as VEGF in tumour-associated endothelial cells and pancreatic cell lines respectively (Karl et al., 2005; Wang et al., 2008). Moreover, (−)-gossypol, a natural BH3 mimetic that inhibits BH3 domain of Bcl-2 as well as related prosurvival proteins (Bcl-xL and Mcl-1), was shown to remarkably decrease microvessel density in human prostate tumour PC-3 xenografts through decrease of VEGF and IL-8 release as well as blocking multiple steps in VEGF-activated biological events (Karaca et al., 2008; Pang et al., 2011).
Matrix degrading and remodelling-targeted therapy
Matrix degrading and remodelling are activated by tumours to modify local micro-environment, which in turn promote their angiogenic potential (Bergers et al., 2000; Vlodavsky and Friedmann, 2001). Up-regulation of expression and activity of several endogenous MMPs including MMP-2, MMP-9 as well as MMP-3 and MMP-7 have been identified in invasive tumours (for a review, see Bourboulia and Stetler-Stevenson, 2010). Consequently, inhibitors of MMPs were extensively pursued as a therapeutic strategy for treating cancer. Unfortunately, MMPs intervention strategies had met with limited clinical success because of severe toxicities and associated metastasis-promoting effect (Coussens et al., 2002; Devy et al., 2009). Furthermore, the paradoxical roles of tissue inhibitors of metalloproteinases (TIMPs) may contribute to such failure depending on the net balance of TIMPs and MMPs in tumour stroma (Jiang et al., 2002). As a result, efforts were directed at therapies exploiting endogenous MMP inhibitors, TIMPs or monoclonal antibodies against individual MMPs (Martens et al., 2007; Jarvelainen et al., 2009). For example, DX-2400, a highly selective fully human MMP-14 inhibitory antibody, was found to block pro-MMP-2 processing on tumour and endothelial cells, inhibited angiogenesis, and slowed tumour progression and formation of metastatic lesions (Devy et al., 2009). Alternatively, in order to reduce toxicity and enhance drug delivery, polymeric nanoparticulate delivery systems could be used to target individual components of ECM. For example, targeted delivery of antisense inhibitors of laminin-8, a vascular basement membrane component, by conjugation to the natural drug carrier β-poly(L-malic acid) significantly reduced tumour microvessel density and increased animal survival in an experimental model of glioblastoma (Fujita et al., 2006). Similarly, a nano delivery system that incorporate peptides against proteolytically processed type IV collagen significantly accumulated in tumours and blocked angiogenesis in experimental models (Mueller et al., 2009). However, the highly sulfated oligosaccharides, Heparan (HS) mimetics highly sulfated oligosaccharides, were shown to have a heparanase-inhibiting effect sequestering, in turn, many heparan sulfate proteoglycan (HSPG)-binding factors (Johnstone et al., 2010; Dredge et al., 2011). In preclinical studies, HS mimetics have effectively targeted multiple HSPG-dependent functions and have resulted in decreased in vivo tumour growth, tumour invasion, tumour metastasis and angiogenesis (Johnstone et al., 2010; Dredge et al., 2011; Zhou et al., 2011). Clinically, the heparanase inhibitor PI-88 showed preliminary efficacy as an adjunct therapy for post-operative hepatocellular carcinoma (Liu et al., 2009).
Tumour-associated stromal cell-targeted therapy
Tumour-associated stromal cells crosstalk is a perquisite for the formation of a tumour vasculature, an essential step for tumour progression (Lorusso and Ruegg, 2008). Interference with those crosstalk circuits through intervention of cellular adhesion (highlighted in next paragraph) or tumour-induced recruitment of different stromal cells may be considered as an indirect way of anti-angiogenic therapy (Ferrara and Kerbel, 2005). The latter can be supported by studies in which inhibition of macrophage infiltration, for example, by either genetic ablation of the macrophage CSF-1 or liposomal clodronate-induced macrophage depletion, was shown to delay the angiogenic switch and malignant transition (Giraudo et al., 2004; Lin et al., 2006). Furthermore, CSF-1R kinase inhibitors were found to reduce tumour-associated vascularity in two different tumour mouse models (Kubota et al., 2009; Manthey et al., 2009). In addition, clodronate and other related bisphosphonates, originally used to treat skeletal complications in patients with tumour-induced osteolysis, were shown to exert potent anti-tumour and anti-angiogenic effects in many other studies (Fournier et al., 2002; Santini et al., 2003; Stathopoulos et al., 2008). Zoledronic acid, a third-generation bisphosphonate, was also found to reduce a number of tumour-associated macrophages and shift their phenotype from M2 to M1, resulting in a reduction in TAM-associated production of VEGF in murine models of spontaneous mammary carcinogenesis and mesothelioma (Coscia et al., 2010; Veltman et al., 2010). Clinically, repeated low-dose therapy with zoledronic acid, which maintains active drug plasma concentration, was able to induce an early remarkable and long-lasting decrease of VEGF levels in patients with cancer (Santini et al., 2007). In another example, inhibition of mobilization of neutrophils, from bone marrow and their infiltration into tumour, using neutralizing anti–prokineticin-2, an antibody against a secreted protein known also as BV8, was shown to impair the initial angiogenic switch in a multistage pancreatic beta cell tumourigenesis model (Shojaei et al., 2008). Furthermore, the neutralizing anti-BV8 was found to prevent myeloid cell-dependent tumour angiogenesis in several xenograft models (Shojaei et al., 2007). Cancer-associated fibroblasts (CAF) can also be targeted with thapsigargin analogue coupled with peptides specific for fibroblast activation protein (FAP), a CAF membrane-bound protease whose catalytic site has access to the peritumoural fluid of the tumour micro-environment. This extracellular activation results in the death of CAFs as well as pericytes and endothelial cells within milieu of different human tumour xenografts (Brennen et al., 2012).
Cell adhesion molecules (CAMs)-targeted therapy
CAMs are cell surface proteins known to be involved in binding with other counter-receptors on adjacent cells or surrounding ECM macromolecules (Aplin et al., 1998). Many CAMs, such as αv-integrins, E-selectin, N-cadherin and VE-cadherin, have been implicated in tumour angiogenesis (Bischoff, 1997; Tei et al., 2002; Nakashima et al., 2003; Weis and Cheresh, 2011). For example, αv-integrins are expressed on surface of endothelial cells and can determine whether cells can adhere to and survive in a particular micro-environment. A number of matrix-derived fragments have the ability to act as endogenous angiogenesis inhibitors through binding to integrins on endothelial cells, disrupting physical connections and suppressing signalling events associated with cell survival, migration and proliferation (Nyberg et al., 2005). Consequently, integrins antagonism using peptidomimetics (e.g. cilengitide), monoclonal antibodies (e.g. volociximab) or oral small-molecule compounds have been investigated in a wide range of malignancies (Huveneers et al., 2007). Cilengitide is a cyclized pentapeptide peptidomimetic designed to compete for the arginine-glycine-aspartic acid (RGD) peptide sequence, thereby blocking the ligation of the αvβ3 and αvβ5 integrins to matrix proteins (Hariharan et al., 2007). Cilengitide is mainly under clinical development for glioblastoma; however, clinical trials of other malignancies such as head and neck cancer as well as lung cancer were also initiated (Reardon and Cheresh, 2011; Vermorken et al., 2012; Manegold et al., 2013). Alternatively, cyclic peptides containing RGD motif could guide nanoparticulate delivery system, which incorporates anti-angiogenic cytotoxic agents such as doxorubicin, paclitaxel or combretastatin A4, to accumulate specifically in tumour vasculature with no overt systemic toxicity (Murphy et al., 2008; Ruoslahti et al., 2010; Wang et al., 2011). Volociximab, a chimeric humanized monoclonal antibody that selectively inhibits the αvβ1 integrin interaction with fibronectin, has been evaluated also in clinical trials for solid tumours such as renal cell carcinoma, recurrent ovarian cancer, advanced non–small-cell lung cancer and metastatic pancreatic cancer (Figlin et al., 2006; Evans et al., 2007; Jarvelainen et al., 2009; Vergote et al., 2009; Besse et al., 2013). Cadherins constitute a superfamily of molecules that mediate calcium-dependent cell–cell adhesions. The intracellular domains of cadherins directly bind to β-catenin and link with cytoskeletal components, providing the molecular basis for stable cell–cell adhesion (Zhang et al., 2010). Targeting cadherin signalling may also represent another way for tumour angiogenesis intervention. For example, ADH-1, a cyclic pentapeptide containing the cell adhesion recognition site (His-Ala-Val) required for N-cadherin adhesion, was shown to possess anti-angiogenic and anti-tumour activity (Blaschuk et al., 2005; Blaschuk, 2012). Similarly, monoclonal antibody directed against specific region of VE-cadherin was able to inhibit tumour angiogenesis and growth with no side effects on normal vasculature (Corada et al., 2002; May et al., 2005).
Inflammatory angiogenesis-targeted therapy
Targeting inflammatory angiogenesis, responsible for a substantial part of tumour vascularization initiated by infiltrating leukocytes, may be considered as another indirect anti-angiogenic strategy (Albini et al., 2005). Moreover, as mentioned before, tumour-infiltrating leukocytes contribute into malignant progression through production of many pro-inflammatory cytokines, chemokines and enzymes that can mostly induce angiogenic cascade (Balkwill et al., 2005). Such vital roles have been supported by the early observation that nonsteroidal anti-inflammatory drugs can inhibit tumour angiogenesis and, in turn, tumour progression (Albini et al., 2005). For example, ibuprofen was found to decrease tumour growth and metastatic potential in mice models through modulation of angiogenesis (Yao et al., 2005). Moreover, selective inhibitors of COX-2, an inducible enzyme that catalyses the production of prostanoids from arachidonic acid, were also shown to inhibit angiogenesis (Tsujii et al., 1998; Wei et al., 2004). The anti-angiogenic effect of COX-2 inhibitors may be contributed, in part, by decreasing the COX-2 metabolic product PGE2, the predominant PG in solid tumours known to stimulate cancer cells to produce pro-angiogenic factors such as VEGF and bFGF as well as many other factors belonging to CXC chemokines family (Strieter et al., 2004; Wang et al., 2006; Wang and Dubois, 2010). Members of the CXC chemokine family are heparin-binding proteins that possess disparate regulative roles in angiogenesis. For example, the ELR+ CXC chemokines, characterized by highly conserved three amino acid motifs (Glu-Leu-Arg; ‘ELR’ motif), are potent promoters of angiogenesis, whereas the IFN-inducible (ELR−) CXC chemokines are inhibitors of angiogenesis (Strieter et al., 2004). The use of repertaxin, originally designed to target the ELR+ CXC chemokine receptors CXCR1 and CXCR2 on neutrophils to prevent their migration to sites of inflammation, was found to inhibit tumour angiogenesis, thereby suppressing tumour progression in a genetic model of pancreatic ductal adenocarcinoma (Ijichi et al., 2011). It would be beneficial to explore other small-molecule CXCR2 antagonists that have already been developed for the treatment of inflammatory diseases in different preclinical models of cancer, especially inflammation-associated cancers (refer to Chapman et al., 2009 for a list of newly developed CXCR2 antagonists used in the treatment of inflammatory diseases of the lung).
Treatment rationales of angiogenesis inhibitors
Angiogenesis inhibitors are used as either monotherapy or in combination with other anti-tumour drugs. Monotherapy using anti-angiogenic agents is mostly intended for prevention of cancer in susceptible individuals or for delaying disease progression in patients with cancer who have previously treated with first-line/second-line regimens. In combined therapy, anti-angiogenic drugs may be added to treatment regimens to increase their efficacy or to reduce developing drug resistance.
Rationale for monotherapy
Patients with cancer who have been treated with first-line or second-line regimens can proceed on a maintenance therapy using one anti-angiogenic drug to extend the progression-free survival (Giuliani et al., 2010; Ledermann et al., 2011; Fabi et al., 2012). Furthermore, anti-angiogenic monotherapy may be beneficial in patients who are at high risk for developing cancer through interference with the angiogenic switch (O'Reilly et al., 1997; Bergers and Benjamin, 2003). This kind of strategy is supported by the comparative safety of anti-angiogenic drugs allowing administration over extended periods (Ma and Waxman, 2008). Many natural and synthetic agents have been shown to significantly reduce the risk of cancer development. Aspirin, a non-selective inhibitor of COX-1 and -2, was associated with a reduced long-term incidence and mortality due to colorectal cancer (Rothwell et al., 2010). The selective COX-2 inhibitor, celecoxib, was also shown to reduce the risk of developing colorectal adenomas in susceptible individuals (Bertagnolli et al., 2006). However, many natural anti-angiogenic molecules present in numerous dietary sources can protect against cancer development and progression (Li et al., 2012). For example, resveratrol, a natural phytoalexin and polyphenol, was found to prevent the onset of colon cancer in a mouse model of colitis-driven colon cancer (Cui et al., 2010). Resveratrol also inhibited chemically induced mammary carcinogenesis in rats as well as tumour growth and metastasis in mice bearing Lewis lung carcinoma (Kimura and Okuda, 2001; Banerjee et al., 2002).
Rationale for combinatory therapy
Crosstalk between angiogenic and oncogenic signalling pathways provides a strong rationale for combining angiogenesis inhibitors with chemotherapeutic and targeted anticancer agents. The consequent overall therapeutic response depends on appropriate designing of the combinatory treatment schedules (Ma and Waxman, 2008). Combination schedule of angiogenesis inhibitors can be neoadjuvant (before the chemotherapeutic drug), concurrent or adjuvant (after the chemotherapeutic drug) depending on the tumour type, anti-angiogenic drug and the chemotherapeutic agent itself (Li et al., 2002; Ma and Waxman, 2009). Using such combinatory rationale has been proved to be more effective than individual treatments in many preclinical and clinical studies. For example, bortezomib, a proteasome inhibitor, was shown to enhance the growth inhibitory effects of IFN-α in the human bladder UM-UC-3 tumours, due to down-regulation of VEGF with ultimate decrease in microvessel densities (Papageorgiou et al., 2006). In another study, combination treatment with vatalanib, a VEGF TKI, and dacinostat, a histone deacetylase inhibitor, was more effective than single agents in inhibiting in vitro and in vivo VEGF-induced angiogenesis, with a synergistic growth inhibitory effect on mouse models of subcutaneous prostate and orthotopic breast tumours (Qian et al., 2004). A significant therapeutic improvement was also achieved when cyclophosphamide was included in the combination therapy with axitinib, another VEGF TKI, in prostate cancer PC-3 xenografts (Ma and Waxman, 2009). Clinically, the addition of bevacizumab to fluorouracil-based combination chemotherapy results in survival enhancement among patients with metastatic colorectal cancer (Hurwitz et al., 2004; Giantonio et al., 2007).
Mechanisms of enhanced therapeutic efficacy
Dual targeting of tumour vasculature
The activity of angiogenesis inhibitors on vascular cells could be potentiated when administered in combination with chemotherapeutic agents that themselves have vascular targeting properties (Naumova et al., 2006). For example, the addition of paclitaxel to SU6668, a potent inhibitor of VEGFR2, FGFR1 and PDGF-β, was shown to inhibit ovarian carcinoma xenograft progression in the peritoneal cavities of nude mice (Garofalo et al., 2003; Klenke et al., 2007). This synergistic effect of paclitaxel may be attributed to its microtubule-binding properties that were known to correlate significantly with its anti-angiogenic and vascular-disrupting properties (Naumova et al., 2006; Schwartz, 2009).
Targeting different cell types of tumour micro-environment
Enhanced therapeutic effect of anti-angiogenic and cytotoxic therapy combinations may be attributed to destruction of two separate compartments of tumours: cancer cells and endothelial cells (Teicher, 1996). The cytotoxic agents would destroy cancer cells directly, and the anti-angiogenic agents would kill cancer cells indirectly by depriving them of nutrients. Moreover, as mentioned before, chemotherapeutic agents may also have anti-angiogenic effects by targeting tumour endothelial cells and endothelial precursors, and thus enhancing the indirect killing of cancer cells (Hicklin and Ellis, 2005; Jain, 2005). Similarly, dual pericytes and endothelial cell targeting was more effective when combinations of PDGFR(s) antagonists with a VEGFR2 inhibitor have been shown experimentally to greatly disturb pericyte–endothelial cell interactions with a resulted tumour regression (Bergers and Benjamin, 2003).
Normalization of tumour vasculature
During angiogenesis, VEGF induces microvascular permeability that increases deposition of fibrin and other plasma proteins in the tumour stroma leading to high interstitial fluid pressure within tumour micro-environment (Nagy et al., 2006). The high interstitial fluid pressure limits chemotherapeutic drug delivery, a major limitation that was found to be ameliorated by co-treatment with angiogenic inhibitors through normalization of tumour vasculature and relieving local tumour oedema (Jain, 2001; Lammerts et al., 2002; Tong et al., 2004). For example, an anti-angiogenic antibody directed against VEGF was found to normalize tumour vasculature, creating an open therapeutic window during which the chemotherapeutic drug can be incorporated with a consequent maximum drug delivery (Tong et al., 2004). To optimize the benefit of vascular normalization-enhanced tumour drug delivery, the duration of the open window during anti-angiogenesis treatment needs to be better defined by improving imaging techniques, which can measure the spatial and temporal changes in blood flow and other physiological parameters with higher resolution (Jain, 2005). An ongoing clinical trial is currently recruiting patients to test whether short course of low-dose sunitinib can normalize tumour vasculature and enhance tumour delivery of docetaxel, and whether that will improve treatment response and progression-free survival in several refractory solid tumours (http://ClinicalTrials.gov identifier: NCT01803503).
Chemosensitization of tumour cells
Co-administration of some other anti-angiogenic agents with conventional chemotherapy was found to increase tumour hypoxia and drug retention rather than vasculature normalization (Franco et al., 2006; Ma and Waxman, 2009; Ma et al., 2011). Although reduction in tumour drug uptake was detected in some studies, overall therapeutic response was found to be enhanced (Ma and Waxman, 2009; Bello et al., 2013). The conflicting results regarding the aforementioned theories may highlight chemosensitization on the molecular level as a mechanism of enhanced therapeutic efficacy. In this context, anti-angiogenic drugs could augment the direct cytotoxic effect of chemotherapeutic drugs by interfering with survival pathways and/or enhancing drug sensitivity. Growing evidence supports chemosensitization as an important mechanism of enhanced therapeutic efficacy. Different types of tumour cells are capable of expressing the potent pro-angiogenic factor VEGF in addition to different types of VEGFRs on their surface, creating a survival autocrine loops that can be targeted directly by anti-angiogenic therapy (Hicklin and Ellis, 2005; Stacchiotti et al., 2011). Moreover, cancer cells tend to overexpress some other RTKs, which can be targeted by dual anti-angiogenic drugs improving tumour response towards chemotherapy (Folkman, 2007). For example, triple-negative breast xenografts completely regressed when E-3810, inhibitor of VEGFR1–3 and FGFR-1 TKs, was combined with paclitaxel (Bello et al., 2013). The addition of imatinib, inhibitor of BCR-ABL TK, c-Kit and PDGFRs, to 5-Fluoruracil or paclitaxel elicited an enhanced cytotoxic effects characterized by induction of apoptotic cell death and inhibition of tumour angiogenesis along with associated down-regulated tumour cell PDGF-BB and PDGFR-β (Kim et al., 2005). Furthermore, dual inhibition of VEGFR and EGFR using vandetanib (ZD6474) resulted in significant tumour growth inhibition in a range of histologically diverse human xenograft models (Wedge et al., 2002; Holden et al., 2005; Yoshikawa et al., 2009). When moved for clinical development, vandetanib demonstrated a high therapeutic efficacy in phases I–III trials against metastatic medullary thyroid cancer, which lead eventually to FDA approval (Leboulleux et al., 2012; Wells et al., 2012; Ton et al., 2013). However, in many other clinical trials, vandetanib showed either acceptable clinical activities along with safety concerns or limited clinical activities that warrant pre-selection of patients (Blackhall et al., 2010; Hsu et al., 2012; Kreisl et al., 2012; Lee et al., 2012; Meyerhardt et al., 2012).
Interference with the repair of cytotoxic drug-induced damage and resistance mechanisms
Anti-angiogenic therapy may interfere with cytotoxic drug/radiotherapy-induced damage repair, a protective mechanism that occur in response to chemotherapy or radiotherapy and considered as a main reason for disease recurrence (Ma and Waxman, 2008). For example, the addition of mTOR inhibitors to radiation therapy could prevent protection mechanisms against radiation-mediated cytotoxicity of tumour blood vessels (Gorski et al., 1999; Shinohara et al., 2005; Manegold et al., 2008). Furthermore, it has been reported that radiotherapy promotes integrin-mediated survival signalling through PKB/Akt by up-regulating αvβ3 expression on endothelial cells. This escape mechanism can be circumvented by administering the αvβ3 integrin antagonist S247, which prevents radiation-induced PKB/Akt phosphorylation leading to enhanced anti-angiogenic and anti-tumour effects (Abdollahi et al., 2005). Clinically, volociximab, an anti-αvβ1 integrin antibody, in combination with carboplatin and paclitaxel showed a preliminary evidence of efficacy in patients with advanced, untreated non–small-cell lung cancer (Besse et al., 2013). Similarly, vascular-disrupting agents that interfere with tumour growth through occlusion of existing blood vessels, such as combretastin-A4 phosphate, can induce acute mobilization and homing of circulating endothelial precursors to tumour as a protective mechanism. Disruption of this circulating endothelial precursors spike by anti-angiogenic drugs or by genetic manipulation resulted in marked reductions in tumour rim size and blood flow as well as enhanced anti-tumour activity of the vascular-disrupting agents (Shaked et al., 2006).
Consequences of anti-angiogenic therapy with other anticancer therapy
Contrary to initial expectations, treatment with angiogenesis inhibitors was associated with unexpected toxicities. The toxicity profiles of those inhibitors reflect the systemic disturbance of growth factor signalling pathways that mediate their anti-angiogenic activity (Elice and Rodeghiero, 2010; 2012). In this context, disturbance of the tight endothelial cell-platelet interaction that maintains vascular integrity results in bleeding complications, gastrointestinal perforations, and disturbed wound and ulcer healing (Verheul and Pinedo, 2007). In general, the incidence of those adverse effects increases when anti-angiogenic agent is combined with chemotherapy. For example, bleeding complications have been observed in patients with colorectal cancer treated with chemotherapy in combination with bevacizumab (Kabbinavar et al., 2003; Giantonio et al., 2006). In non–small-cell lung cancer, some patients treated with bevacizumab in combination with carboplatin and paclitaxel experienced severe or fatal pulmonary haemorrhage (Johnson et al., 2004). Furthermore, a higher incidence of gastrointestinal perforation was observed in patients with colorectal cancer given bevacizumab in combination with chemotherapy compared with chemotherapy alone (Hurwitz et al., 2004). Similarly, thrombotic events have been observed in patients treated with angiogenesis inhibitors, especially when these agents are given in combination with chemotherapy (Verheul and Pinedo, 2007). Treatment of patients with cancer with angiogenesis inhibitors is frequently associated with hypertension, which may require the addition of regular anti-hypertensive agent (Izzedine et al., 2009). However, patients on anti-angiogenic therapy can develop resistance either via classical resistance mechanisms, such as increased drug metabolism or an increased number of drug efflux pumps, or via compensatory release of different angiogenic inducers (Kerbel and Folkman, 2002; Verheul and Pinedo, 2007). For example, VEGF-targeted therapies were shown to produce a period of stable disease followed by VEGF independent vascular regrowth, promoted cell invasiveness and metastasis (Ebos et al., 2009; Loges et al., 2010). Such resistance was thought to be mediated in part by pro-angiogenic ligands of the FGF family as well as BV8 secretion associated with enhanced recruitment of myeloid cells to hypoxic tumours (Casanovas et al., 2005; Shojaei et al., 2007; Loges et al., 2010). Additionally, anti-VEGF refractoriness may be conferred to CAF-mediated release of PDGF-C that can direct the process of endothelial cell migration and angiogenesis, independent of VEGF as well as that can increase pericytes recruitment and vessels stabilizing minimizing their dependence on VEGF for survival (Crawford et al., 2009; di Tomaso et al., 2009). Consequently, targeting those angiogenesis signalling pathways may lead to enhanced response in anti-VEGF-resistant tumours (Hsu and Wakelee, 2009). Another possibility shown to circumvent this resistance scenario was to combine the VEGFR2 inhibitor cediranib or the monoclonal antibody bevacizumab with cilengitide, the integrin inhibitor (Reardon and Cheresh, 2011).
Summary and future directions
Angiogenesis is a critical process that occurs pathologically in many malignancies due to changing balance of endogenous angiogenesis inducers and inhibitors, leading to the activation of nearby endothelial cells to form new vasculature. Consequently, angiogenesis can be targeted to restrict initiation, growth and progression of most of angiogenesis-dependent malignancies. Numerous angiogenic inhibitors have been identified, some of which are currently being investigated in clinical trials and some others were even approved for cancer therapies. These angiogenesis inhibitors were classified based on their target into two main classes: direct and indirect inhibitors. Indirect angiogenesis inhibitors can be further subclassified based on their interference mechanisms with the angiogenic cascade. A list of major categories and molecular targets for angiogenesis inhibitors is shown in Table 2.
Table 2.
Major categories of angiogenesis inhibitors and molecular targets
| Category of angiogenesis inhibitors | Examples | Target/Mechanism | References |
|---|---|---|---|
| Chemotherapeutic agents | Cyclophosphamide | Induces EC apoptosis, decreases circulating EPC | Hamano et al., 2004; Shahrzad et al., 2008 |
| Paclitaxel | Microtubule | Naumova et al., 2006; Schwartz, 2009 | |
| VEGF-targeted therapy | Bevacizumab | VEGF-A | Hsu and Wakelee, 2009 |
| VEGF-Trap | VEGF-A, VEGF-B and PlGF | Rudge et al., 2007 | |
| Sunitinib | VEGFR1–3, PDGFR-α, PDGFR-β, c-Kit, CSF-1R and Flt-3 | Huang et al., 2010 | |
| Sorafenib | VEGFR1–3, PDGFR-β, Raf-1, B-Raf | Huang et al., 2010 | |
| Vatalanib | VEGFR1–3, PDGFR-β and c-Kit | Qian et al., 2004; Jubb et al., 2006 | |
| Axitinib | VEGFRs, PDGFR-β and c-Kit | Jubb et al., 2006; Ma and Waxman, 2009 | |
| SU6668 | VEGFR2, FGFR1 and PDGF-β | Klenke et al., 2007 | |
| FGF-targeted therapy | AZD4547 | FGFR1–3 | Gavine et al., 2012 |
| Ponatinib | FGFR1–4 | Gozgit et al., 2012 | |
| SSR | FGFRs | Bono et al., 2013 | |
| Brivanib | VEGFRs and FGFRs | Allen et al., 2011 | |
| Dovitinib | FEGFRs, VEGFRs and PDGFR | Chen et al., 2012 | |
| Nintedanib | VEGFRs, FGFRs and PDGFR | Hilberg et al., 2008 | |
| Oncogene-targeted therapy/signalling transduction-targeted therapy | Dasatinib | Src and indirectly VEGF, IL-8 | Summy et al., 2005; Han et al., 2006; Haura et al., 2010 |
| Tipifarnib | MMP-1 | Izbicka et al., 2005 | |
| NVP-AUY922 | Hsp90 | Eccles et al., 2008; Moser et al., 2012 | |
| Bortezomib | NF-κB-dependent release of VEGF and IL-8 | Sunwoo et al., 2001 | |
| Gossypol | VEGF and IL-8 release | Pang et al., 2011 | |
| Dacinostat | Histone deacetylase | Qian et al., 2004 | |
| Matrix degrading and remodelling-targeted therapy | DX-2400 | MMP-14 | Devy et al., 2009 |
| PI-88 | Heparanase | Liu et al., 2009 | |
| Tumour-associated stromal cell-targeted therapy | JNJ-28312141 | CSF-1R | Manthey et al., 2009 |
| Zoledronic acid | TAM-associated production of VEGF | Coscia et al., 2010; Veltman et al., 2010 | |
| Anti-BV8 antibody | Neutrophils recruitment | Shojaei et al., 2008 | |
| CAMs-targeted therapy | Cilengitide | αvβ3 and αvβ5 integrins ligation to matrix proteins | Hariharan et al., 2007 |
| Volociximab | αvβ1 integrin interaction with fibronectin | Evans et al., 2007; Besse et al., 2013 | |
| ADH-1 | N-cadherin | Blaschuk et al., 2005; Blaschuk, 2012 | |
| Inflammatory angiogenesis-targeted therapy | Ibuprofen | COX1/2 | Yao et al., 2005 |
| Celecoxib | COX-2 | Wei et al., 2004 | |
| Repertaxin | CXCR1 and CXCR2 | Ijichi et al., 2011 |
EC, endothelial cells; EPC, endothelial progenitor cells; Hsp90, heat shock protein 90; PlGF, placental growth factor; Src, sarcoma.
Most angiogenesis inhibitors conferred clinical benefits mainly when combined with other chemotherapeutic/targeted therapies rather than being used as monotherapy. Unfortunately, many anti-angiogenic agents were shown to be associated with overt systemic toxicity as well as resistance emergence and disease recurrence. Drug resistance in anti-angiogenic therapy may result from a plethora of pro-angiogenic factors released by inappropriately functioning host cells in the tumour micro-environment as a compensatory mechanism. Therefore, the strategy of targeting endothelial cells alone may not be enough as explained in the previous texts, requiring the proposal of different rationales in which other cellular compartments of tumour micro-environment are targeted to attain proper anti-angiogenic and anti-tumour response. That highlights the importance of considering tumour micro-environment as a dynamic system, as depicted in Figure 1 in which interference with any of its components may be an approach to interfere with cancer hallmarks, including angiogenesis. Meanwhile, differentially expressed molecules on various types of tumour-associated stromal cells can be used as docking sites to concentrate drug conjugates and nanoparticles. That high local concentration of anti-angiogenic agents in tumour micro-environment may lead to increased sustained biological activities and/or reduced adverse effects. In conclusion, future anti-angiogenic treatment(s) should be designed as targeted delivery systems with the ability to accumulate selectively in tumours and/or an ability to shift micro-environmental equilibria towards tumour-unfavourable conditions. The consequent sustained disturbance of dynamic tumour micro-environment may lead eventually to tumour regression, hopefully without systemic toxicity and notable activation of resistance mechanisms.
Figure 1.
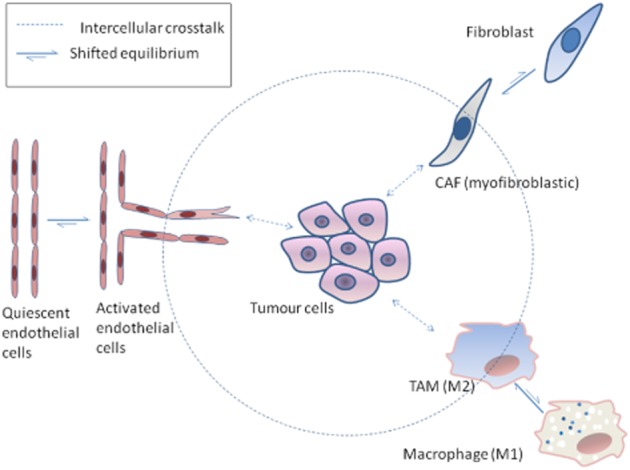
A representation of some cellular components of tumour micro-environment with shifted equilibria towards tumour-favouring phenotypes [CAFs with myofibroblastic characteristics; TAMs with M2 phenotype shift to M1 phenotype, a pro-inflammatory phenotype with a tumouricidal activity.
Acknowledgments
The authors want to acknowledge Dr. Hassan A. El-Kashef, Professor of Pharmacology at Faculty of Pharmacy, Mansoura University, for suggesting the topic of the review. Source of funding: RO1-EY-022408 to A. B. E.
Glossary
- ANG2
angiopoietin-2
- BV8
prokineticin-2
- CAFs
carcinoma-associated fibroblasts
- CAM
cell adhesion molecules
- CSF-1
colony-stimulating factor-1
- ECM
extracellular matrix
- EGFR
EGF receptor
- FGF
fibroblast growth factor
- HIF-1α
hypoxia inducible factor-1α
- HSPG
heparan sulfate proteoglycan
- RTKs
receptor TKs
- S1P
sphingosine-1-phosphate
- TAMs
tumour-associated macrophages
- TKIs
TK inhibitors
Conflict of interest
The authors declare that they have no conflict of interest.
References
- Abdollahi A, Hahnfeldt P, Maercker C, Grone HJ, Debus J, Ansorge W, et al. Endostatin's antiangiogenic signaling network. Mol Cell. 2004;13:649–663. doi: 10.1016/s1097-2765(04)00102-9. [DOI] [PubMed] [Google Scholar]
- Abdollahi A, Griggs DW, Zieher H, Roth A, Lipson KE, Saffrich R, et al. Inhibition of alpha(v)beta3 integrin survival signaling enhances antiangiogenic and antitumor effects of radiotherapy. Clin Cancer Res. 2005;11:6270–6279. doi: 10.1158/1078-0432.CCR-04-1223. [DOI] [PubMed] [Google Scholar]
- Albini A, Tosetti F, Benelli R, Noonan DM. Tumor inflammatory angiogenesis and its chemoprevention. Cancer Res. 2005;65:10637–10641. doi: 10.1158/0008-5472.CAN-05-3473. [DOI] [PubMed] [Google Scholar]
- Ali TK, El-Remessy AB. Diabetic retinopathy: current management and experimental therapeutic targets. Pharmacotherapy. 2009;29:182–192. doi: 10.1592/phco.29.2.182. [DOI] [PubMed] [Google Scholar]
- Allen E, Walters IB, Hanahan D. Brivanib, a dual FGF/VEGF inhibitor, is active both first and second line against mouse pancreatic neuroendocrine tumors developing adaptive/evasive resistance to VEGF inhibition. Clin Cancer Res. 2011;17:5299–5310. doi: 10.1158/1078-0432.CCR-10-2847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Angevin E, Lopez-Martin JA, Lin C-C, Gschwend E, Jr, Harzstark A, Castellano D, et al. Phase I study of dovitinib (TKI258), an oral FGFR, VEGFR, and PDGFR inhibitor, in advanced or metastatic renal cell carcinoma. Clin Cancer Res. 2013;19:1257–1268. doi: 10.1158/1078-0432.CCR-12-2885. [DOI] [PubMed] [Google Scholar]
- Aplin AE, Howe A, Alahari SK, Juliano RL. Signal transduction and signal modulation by cell adhesion receptors: the role of integrins, cadherins, immunoglobulin-cell adhesion molecules, and selectins. Pharmacol Rev. 1998;50:197–263. [PubMed] [Google Scholar]
- Arany Z, Foo SY, Ma Y, Ruas JL, Bommi-Reddy A, Girnun G, et al. HIF-independent regulation of VEGF and angiogenesis by the transcriptional coactivator PGC-1alpha. Nature. 2008;451:1008–1012. doi: 10.1038/nature06613. [DOI] [PubMed] [Google Scholar]
- Argiris A, Karamouzis MV, Gooding WE, Branstetter BF, Zhong S, Raez LE, et al. Phase II trial of pemetrexed and bevacizumab in patients with recurrent or metastatic head and neck cancer. J Clin Oncol. 2011;29:1140–1145. doi: 10.1200/JCO.2010.33.3591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Argiris A, Kotsakis AP, Hoang T, Worden FP, Savvides P, Gibson MK, et al. Cetuximab and bevacizumab: preclinical data and phase II trial in recurrent or metastatic squamous cell carcinoma of the head and neck. Ann Oncol. 2013;24:220–225. doi: 10.1093/annonc/mds245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Awada A, Eskens FA, Piccart M, Cutler DL, van der Gaast A, Bleiberg H, et al. Phase I and pharmacological study of the oral farnesyl transferase inhibitor SCH 66336 given once daily to patients with advanced solid tumours. Eur J Cancer. 2002;38:2272–2278. doi: 10.1016/s0959-8049(02)00379-9. [DOI] [PubMed] [Google Scholar]
- Baeriswyl V, Christofori G. The angiogenic switch in carcinogenesis. Semin Cancer Biol. 2009;19:329–337. doi: 10.1016/j.semcancer.2009.05.003. [DOI] [PubMed] [Google Scholar]
- Baka S, Clamp AR, Jayson GC. A review of the latest clinical compounds to inhibit VEGF in pathological angiogenesis. Expert Opin Ther Targets. 2006;10:867–876. doi: 10.1517/14728222.10.6.867. [DOI] [PubMed] [Google Scholar]
- Balkwill F, Charles KA, Mantovani A. Smoldering and polarized inflammation in the initiation and promotion of malignant disease. Cancer Cell. 2005;7:211–217. doi: 10.1016/j.ccr.2005.02.013. [DOI] [PubMed] [Google Scholar]
- Banerjee S, Bueso-Ramos C, Aggarwal BB. Suppression of 7,12-dimethylbenz(a)anthracene-induced mammary carcinogenesis in rats by resveratrol: role of nuclear factor-kappaB, cyclooxygenase 2, and matrix metalloprotease 9. Cancer Res. 2002;62:4945–4954. [PubMed] [Google Scholar]
- Bello E, Taraboletti G, Colella G, Zucchetti M, Forestieri D, Licandro SA, et al. The tyrosine kinase inhibitor E-3810 combined with paclitaxel inhibits the growth of advanced-stage triple-negative breast cancer xenografts. Mol Cancer Ther. 2013;12:131–140. doi: 10.1158/1535-7163.MCT-12-0275-T. [DOI] [PubMed] [Google Scholar]
- Bergers G, Benjamin LE. Tumorigenesis and the angiogenic switch. Nat Rev Cancer. 2003;3:401–410. doi: 10.1038/nrc1093. [DOI] [PubMed] [Google Scholar]
- Bergers G, Hanahan D. Modes of resistance to anti-angiogenic therapy. Nat Rev Cancer. 2008;8:592–603. doi: 10.1038/nrc2442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergers G, Brekken R, McMahon G, Vu TH, Itoh T, Tamaki K, et al. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nat Cell Biol. 2000;2:737–744. doi: 10.1038/35036374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertagnolli MM, Eagle CJ, Zauber AG, Redston M, Solomon SD, Kim K, et al. Celecoxib for the prevention of sporadic colorectal adenomas. N Engl J Med. 2006;355:873–884. doi: 10.1056/NEJMoa061355. [DOI] [PubMed] [Google Scholar]
- Besse B, Tsao LC, Chao DT, Fang Y, Soria JC, Almokadem S, et al. Phase Ib safety and pharmacokinetic study of volociximab, an anti-alpha5beta1 integrin antibody, in combination with carboplatin and paclitaxel in advanced non-small-cell lung cancer. Ann Oncol. 2013;24:90–96. doi: 10.1093/annonc/mds281. [DOI] [PubMed] [Google Scholar]
- Bid HK, Zhan J, Phelps DA, Kurmasheva RT, Houghton PJ. Potent inhibition of angiogenesis by the IGF-1 receptor-targeting antibody SCH717454 is reversed by IGF-2. Mol Cancer Ther. 2011;11:649–659. doi: 10.1158/1535-7163.MCT-11-0575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bischoff J. Cell adhesion and angiogenesis. J Clin Invest. 1997;100(Suppl. 11):S37–S39. [PubMed] [Google Scholar]
- Blackhall FH, O'Brien M, Schmid P, Nicolson M, Taylor P, Milenkova T, et al. A phase I study of Vandetanib in combination with vinorelbine/cisplatin or gemcitabine/cisplatin as first-line treatment for advanced non-small cell lung cancer. J Thorac Oncol. 2010;5:1285–1288. doi: 10.1097/JTO.0b013e3181e3a2d1. [DOI] [PubMed] [Google Scholar]
- Blaschuk O, Lavoie N, Devamy E, Lepekhin E. Further observations concerning the effects of the N-cadherin antagonist ExherinTM (ADH-1) on endothelial cells and tumor blood vessels. AACR Meeting Abstracts. 2005;2005(1):268. [Google Scholar]
- Blaschuk OW. Discovery and development of N-cadherin antagonists. Cell Tissue Res. 2012;348:309–313. doi: 10.1007/s00441-011-1320-5. [DOI] [PubMed] [Google Scholar]
- Bono F, De Smet F, Herbert C, De Bock K, Georgiadou M, Fons P, et al. Inhibition of tumor angiogenesis and growth by a small-molecule multi-FGF receptor blocker with allosteric properties. Cancer Cell. 2013;23:477–488. doi: 10.1016/j.ccr.2013.02.019. [DOI] [PubMed] [Google Scholar]
- Bouis D, Kusumanto Y, Meijer C, Mulder NH, Hospers GAP. A review on pro- and anti-angiogenic factors as targets of clinical intervention. Pharmacol Res. 2006;53:89–103. doi: 10.1016/j.phrs.2005.10.006. [DOI] [PubMed] [Google Scholar]
- Bourboulia D, Stetler-Stevenson WG. Matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs): positive and negative regulators in tumor cell adhesion. Semin Cancer Biol. 2010;20:161–168. doi: 10.1016/j.semcancer.2010.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brennen WN, Rosen DM, Wang H, Isaacs JT, Denmeade SR. Targeting carcinoma-associated fibroblasts within the tumor stroma with a fibroblast activation protein-activated prodrug. J Natl Cancer Inst. 2012;104:1320–1334. doi: 10.1093/jnci/djs336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Briasoulis E, Aravantinos G, Kouvatseas G, Pappas P, Biziota E, Sainis I, et al. Dose selection trial of metronomic oral vinorelbine monotherapy in patients with metastatic cancer: a hellenic cooperative oncology group clinical translational study. BMC Cancer. 2013;13:263. doi: 10.1186/1471-2407-13-263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burger RA, Brady MF, Bookman MA, Fleming GF, Monk BJ, Huang H, et al. Incorporation of bevacizumab in the primary treatment of ovarian cancer. N Engl J Med. 2011;365:2473–2483. doi: 10.1056/NEJMoa1104390. [DOI] [PubMed] [Google Scholar]
- Burstein HJ. Bevacizumab for advanced breast cancer: all tied up with a RIBBON? J Clin Oncol. 2011;29:1232–1235. doi: 10.1200/JCO.2010.33.2684. [DOI] [PubMed] [Google Scholar]
- Carmeliet P. Angiogenesis in health and disease. Nat Med. 2003;9:653–660. doi: 10.1038/nm0603-653. [DOI] [PubMed] [Google Scholar]
- Carmeliet P. Angiogenesis in life, disease and medicine. Nature. 2005;438:932–936. doi: 10.1038/nature04478. [DOI] [PubMed] [Google Scholar]
- Casanovas O, Hicklin DJ, Bergers G, Hanahan D. Drug resistance by evasion of antiangiogenic targeting of VEGF signaling in late-stage pancreatic islet tumors. Cancer Cell. 2005;8:299–309. doi: 10.1016/j.ccr.2005.09.005. [DOI] [PubMed] [Google Scholar]
- Chapman RW, Phillips JE, Hipkin RW, Curran AK, Lundell D, Fine JS. CXCR2 antagonists for the treatment of pulmonary disease. Pharmacol Ther. 2009;121:55–68. doi: 10.1016/j.pharmthera.2008.10.005. [DOI] [PubMed] [Google Scholar]
- Chen ZY, Shi M, Peng LX, Wei W, Li XJ, Guo ZX, et al. Dovitinib preferentially targets endothelial cells rather than cancer cells for the inhibition of hepatocellular carcinoma growth and metastasis. J Transl Med. 2012;10:245. doi: 10.1186/1479-5876-10-245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu J, Yau T. Metastatic pancreatic cancer: are we making progress in treatment? Gastroenterol Res Pract. 2012;2012:898931. doi: 10.1155/2012/898931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung AS, Lee J, Ferrara N. Targeting the tumour vasculature: insights from physiological angiogenesis. Nat Rev Cancer. 2010;10:505–514. doi: 10.1038/nrc2868. [DOI] [PubMed] [Google Scholar]
- Cirri P, Chiarugi P. Cancer associated fibroblasts: the dark side of the coin. Am J Cancer Res. 2011;1:482–497. [PMC free article] [PubMed] [Google Scholar]
- Cohen MH, Gootenberg J, Keegan P, Pazdur R. FDA drug approval summary: bevacizumab plus FOLFOX4 as second-line treatment of colorectal cancer. Oncologist. 2007;12:356–361. doi: 10.1634/theoncologist.12-3-356. [DOI] [PubMed] [Google Scholar]
- Corada M, Zanetta L, Orsenigo F, Breviario F, Lampugnani MG, Bernasconi S, et al. A monoclonal antibody to vascular endothelial-cadherin inhibits tumor angiogenesis without side effects on endothelial permeability. Blood. 2002;100:905–911. doi: 10.1182/blood.v100.3.905. [DOI] [PubMed] [Google Scholar]
- Coscia M, Quaglino E, Iezzi M, Curcio C, Pantaleoni F, Riganti C, et al. Zoledronic acid repolarizes tumour-associated macrophages and inhibits mammary carcinogenesis by targeting the mevalonate pathway. J Cell Mol Med. 2010;14:2803–2815. doi: 10.1111/j.1582-4934.2009.00926.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coussens LM, Fingleton B, Matrisian LM. Matrix metalloproteinase inhibitors and cancer: trials and tribulations. Science. 2002;295:2387–2392. doi: 10.1126/science.1067100. [DOI] [PubMed] [Google Scholar]
- Crawford Y, Kasman I, Yu L, Zhong C, Wu X, Modrusan Z, et al. PDGF-C mediates the angiogenic and tumorigenic properties of fibroblasts associated with tumors refractory to anti-VEGF treatment. Cancer Cell. 2009;15:21–34. doi: 10.1016/j.ccr.2008.12.004. [DOI] [PubMed] [Google Scholar]
- Cui X, Jin Y, Hofseth AB, Pena E, Habiger J, Chumanevich A, et al. Resveratrol suppresses colitis and colon cancer associated with colitis. Cancer Prev Res (Phila) 2010;3:549–559. doi: 10.1158/1940-6207.CAPR-09-0117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devy L, Huang L, Naa L, Yanamandra N, Pieters H, Frans N, et al. Selective inhibition of matrix metalloproteinase-14 blocks tumor growth, invasion, and angiogenesis. Cancer Res. 2009;69:1517–1526. doi: 10.1158/0008-5472.CAN-08-3255. [DOI] [PubMed] [Google Scholar]
- Dredge K, Hammond E, Handley P, Gonda TJ, Smith MT, Vincent C, et al. PG545, a dual heparanase and angiogenesis inhibitor, induces potent anti-tumour and anti-metastatic efficacy in preclinical models. Br J Cancer. 2011;104:635–642. doi: 10.1038/bjc.2011.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drevs J, Fakler J, Eisele S, Medinger M, Bing G, Esser N, et al. Antiangiogenic potency of various chemotherapeutic drugs for metronomic chemotherapy. Anticancer Res. 2004;24:1759–1763. [PubMed] [Google Scholar]
- Dutt A, Ramos AH, Hammerman PS, Mermel C, Cho J, Sharifnia T, et al. Inhibitor-sensitive FGFR1 amplification in human non-small cell lung cancer. PLoS One. 2011;6:e20351. doi: 10.1371/journal.pone.0020351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebos JM, Lee CR, Cruz-Munoz W, Bjarnason GA, Christensen JG, Kerbel RS. Accelerated metastasis after short-term treatment with a potent inhibitor of tumor angiogenesis. Cancer Cell. 2009;15:232–239. doi: 10.1016/j.ccr.2009.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eccles SA, Massey A, Raynaud FI, Sharp SY, Box G, Valenti M, et al. NVP-AUY922: a novel heat shock protein 90 inhibitor active against xenograft tumor growth, angiogenesis, and metastasis. Cancer Res. 2008;68:2850–2860. doi: 10.1158/0008-5472.CAN-07-5256. [DOI] [PubMed] [Google Scholar]
- Eikesdal HP, Sugimoto H, Birrane G, Maeshima Y, Cooke VG, Kieran M, et al. Identification of amino acids essential for the antiangiogenic activity of tumstatin and its use in combination antitumor activity. Proc Natl Acad Sci U S A. 2008;105:15040–15045. doi: 10.1073/pnas.0807055105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elice F, Rodeghiero F. Bleeding complications of antiangiogenic therapy: pathogenetic mechanisms and clinical impact. Thromb Res. 2010;125(Suppl. 2):S55–S57. doi: 10.1016/S0049-3848(10)70014-1. [DOI] [PubMed] [Google Scholar]
- Elice F, Rodeghiero F. Side effects of anti-angiogenic drugs. Thromb Res. 2012;129(Suppl. 1):S50–S53. doi: 10.1016/S0049-3848(12)70016-6. [DOI] [PubMed] [Google Scholar]
- Ellis LM, Hicklin DJ. VEGF-targeted therapy: mechanisms of anti-tumour activity. Nat Rev Cancer. 2008;8:579–591. doi: 10.1038/nrc2403. [DOI] [PubMed] [Google Scholar]
- El-Remessy AB, Al-Shabrawey M, Platt DH, Bartoli M, Behzadian MA, Ghaly N, et al. Peroxynitrite mediates VEGF's angiogenic signal and function via a nitration-independent mechanism in endothelial cells. FASEB J. 2007;21:2528–2539. doi: 10.1096/fj.06-7854com. [DOI] [PubMed] [Google Scholar]
- Evans T, Ramanathan RK, Yazji S, Glynne-Jones R, Anthoney A, Berlin J, et al. Final results from cohort 1 of a phase II study of volociximab, an anti-{alpha}5{beta}1 integrin antibody, in combination with gemcitabine (GEM) in patients (pts) with metastatic pancreatic cancer (MPC) ASCO Meeting Abstracts. 2007;25(18 Suppl.):4549. [Google Scholar]
- Fabi A, Russillo M, Ferretti G, Metro G, Nistico C, Papaldo P, et al. Maintenance bevacizumab beyond first-line paclitaxel plus bevacizumab in patients with Her2-negative hormone receptor-positive metastatic breast cancer: efficacy in combination with hormonal therapy. BMC Cancer. 2012;12:482. doi: 10.1186/1471-2407-12-482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fagiani E, Christofori G. Angiopoietins in angiogenesis. Cancer Lett. 2013;328:18–26. doi: 10.1016/j.canlet.2012.08.018. [DOI] [PubMed] [Google Scholar]
- Fang P, Hu JH, Cheng ZG, Liu ZF, Wang JL, Jiao SC. Efficacy and safety of bevacizumab for the treatment of advanced hepatocellular carcinoma: a systematic review of phase II trials. PLoS One. 2012;7:e49717. doi: 10.1371/journal.pone.0049717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrara N, Kerbel RS. Angiogenesis as a therapeutic target. Nature. 2005;438:967–974. doi: 10.1038/nature04483. [DOI] [PubMed] [Google Scholar]
- Figlin RA, Kondagunta GV, Yazji S, Motzer RJ, Bukowski RM. Phase II study of volociximab (M200), an {alpha}5{beta}1 anti-integrin antibody in refractory metastatic clear cell renal cell cancer (RCC) ASCO Meeting Abstracts. 2006;24(18 Suppl.):4535. [Google Scholar]
- Folkman J. Tumor angiogenesis: therapeutic implications. N Engl J Med. 1971;285:1182–1186. doi: 10.1056/NEJM197111182852108. [DOI] [PubMed] [Google Scholar]
- Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat Med. 1995;1:27–30. doi: 10.1038/nm0195-27. [DOI] [PubMed] [Google Scholar]
- Folkman J. Angiogenesis and apoptosis. Semin Cancer Biol. 2003;13:159–167. doi: 10.1016/s1044-579x(02)00133-5. [DOI] [PubMed] [Google Scholar]
- Folkman J. Angiogenesis: an organizing principle for drug discovery? Nat Rev Drug Discov. 2007;6:273–286. doi: 10.1038/nrd2115. [DOI] [PubMed] [Google Scholar]
- Fontana A, Bocci G, Galli L, D'Arcangelo M, Derosa L, Fioravanti A, et al. Metronomic cyclophosphamide in elderly patients with advanced, castration-resistant prostate cancer. J Am Geriatr Soc. 2010;58:986–988. doi: 10.1111/j.1532-5415.2010.02833.x. [DOI] [PubMed] [Google Scholar]
- Fournier P, Boissier S, Filleur S, Guglielmi J, Cabon F, Colombel M, et al. Bisphosphonates inhibit angiogenesis in vitro and testosterone-stimulated vascular regrowth in the ventral prostate in castrated rats. Cancer Res. 2002;62:6538–6544. [PubMed] [Google Scholar]
- Franco M, Man S, Chen L, Emmenegger U, Shaked Y, Cheung AM, et al. Targeted anti-vascular endothelial growth factor receptor-2 therapy leads to short-term and long-term impairment of vascular function and increase in tumor hypoxia. Cancer Res. 2006;66:3639–3648. doi: 10.1158/0008-5472.CAN-05-3295. [DOI] [PubMed] [Google Scholar]
- Fujita M, Khazenzon NM, Ljubimov AV, Lee BS, Virtanen I, Holler E, et al. Inhibition of laminin-8 in vivo using a novel poly(malic acid)-based carrier reduces glioma angiogenesis. Angiogenesis. 2006;9:183–191. doi: 10.1007/s10456-006-9046-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garofalo A, Naumova E, Manenti L, Ghilardi C, Ghisleni G, Caniatti M, et al. The combination of the tyrosine kinase receptor inhibitor SU6668 with paclitaxel affects ascites formation and tumor spread in ovarian carcinoma xenografts growing orthotopically. Clin Cancer Res. 2003;9:3476–3485. [PubMed] [Google Scholar]
- Gavine PR, Mooney L, Kilgour E, Thomas AP, Al-Kadhimi K, Beck S, et al. AZD4547: an orally bioavailable, potent, and selective inhibitor of the fibroblast growth factor receptor tyrosine kinase family. Cancer Res. 2012;72:2045–2056. doi: 10.1158/0008-5472.CAN-11-3034. [DOI] [PubMed] [Google Scholar]
- Gaya A, Tse V. A preclinical and clinical review of aflibercept for the management of cancer. Cancer Treat Rev. 2012;38:484–493. doi: 10.1016/j.ctrv.2011.12.008. [DOI] [PubMed] [Google Scholar]
- Gebbia V, Boussen H, Valerio MR. Oral metronomic cyclophosphamide with and without methotrexate as palliative treatment for patients with metastatic breast carcinoma. Anticancer Res. 2012;32:529–536. [PubMed] [Google Scholar]
- Gerber PA, Hippe A, Buhren BA, Muller A, Homey B. Chemokines in tumor-associated angiogenesis. Biol Chem. 2009;390:1213–1223. doi: 10.1515/BC.2009.144. [DOI] [PubMed] [Google Scholar]
- Giannoni E, Bianchini F, Masieri L, Serni S, Torre E, Calorini L, et al. Reciprocal activation of prostate cancer cells and cancer-associated fibroblasts stimulates epithelial-mesenchymal transition and cancer stemness. Cancer Res. 2010;70:6945–6956. doi: 10.1158/0008-5472.CAN-10-0785. [DOI] [PubMed] [Google Scholar]
- Giantonio BJ, Levy DE, O'Dwyer PJ, Meropol NJ, Catalano PJ, Benson AB., 3rd A phase II study of high-dose bevacizumab in combination with irinotecan, 5-fluorouracil, leucovorin, as initial therapy for advanced colorectal cancer: results from the Eastern Cooperative Oncology Group study E2200. Ann Oncol. 2006;17:1399–1403. doi: 10.1093/annonc/mdl161. [DOI] [PubMed] [Google Scholar]
- Giantonio BJ, Catalano PJ, Meropol NJ, O'Dwyer PJ, Mitchell EP, Alberts SR, et al. Bevacizumab in combination with oxaliplatin, fluorouracil, and leucovorin (FOLFOX4) for previously treated metastatic colorectal cancer: results from the Eastern Cooperative Oncology Group Study E3200. J Clin Oncol. 2007;25:1539–1544. doi: 10.1200/JCO.2006.09.6305. [DOI] [PubMed] [Google Scholar]
- Giraudo E, Inoue M, Hanahan D. An amino-bisphosphonate targets MMP-9-expressing macrophages and angiogenesis to impair cervical carcinogenesis. J Clin Invest. 2004;114:623–633. doi: 10.1172/JCI22087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giuliani F, De Vita F, Colucci G, Pisconti S. Maintenance therapy in colon cancer. Cancer Treat Rev. 2010;36(Suppl. 3):S42–S45. doi: 10.1016/S0305-7372(10)70019-0. [DOI] [PubMed] [Google Scholar]
- Gorski DH, Beckett MA, Jaskowiak NT, Calvin DP, Mauceri HJ, Salloum RM, et al. Blockage of the vascular endothelial growth factor stress response increases the antitumor effects of ionizing radiation. Cancer Res. 1999;59:3374–3378. [PubMed] [Google Scholar]
- Gozgit JM, Wong MJ, Moran L, Wardwell S, Mohemmad QK, Narasimhan NI, et al. Ponatinib (AP24534), a multitargeted pan-FGFR inhibitor with activity in multiple FGFR-amplified or mutated cancer models. Mol Cancer Ther. 2012;11:690–699. doi: 10.1158/1535-7163.MCT-11-0450. [DOI] [PubMed] [Google Scholar]
- Hamano Y, Sugimoto H, Soubasakos MA, Kieran M, Olsen BR, Lawler J, et al. Thrombospondin-1 associated with tumor microenvironment contributes to low-dose cyclophosphamide-mediated endothelial cell apoptosis and tumor growth suppression. Cancer Res. 2004;64:1570–1574. doi: 10.1158/0008-5472.can-03-3126. [DOI] [PubMed] [Google Scholar]
- Han JY, Oh SH, Morgillo F, Myers JN, Kim E, Hong WK, et al. Hypoxia-inducible factor 1alpha and antiangiogenic activity of farnesyltransferase inhibitor SCH66336 in human aerodigestive tract cancer. J Natl Cancer Inst. 2005;97:1272–1286. doi: 10.1093/jnci/dji251. [DOI] [PubMed] [Google Scholar]
- Han LY, Landen CN, Trevino JG, Halder J, Lin YG, Kamat AA, et al. Antiangiogenic and antitumor effects of SRC inhibition in ovarian carcinoma. Cancer Res. 2006;66:8633–8639. doi: 10.1158/0008-5472.CAN-06-1410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanahan D, Folkman J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell. 1996;86:353–364. doi: 10.1016/s0092-8674(00)80108-7. [DOI] [PubMed] [Google Scholar]
- Hariharan S, Gustafson D, Holden S, McConkey D, Davis D, Morrow M, et al. Assessment of the biological and pharmacological effects of the alpha nu beta3 and alpha nu beta5 integrin receptor antagonist, cilengitide (EMD 121974), in patients with advanced solid tumors. Ann Oncol. 2007;18:1400–1407. doi: 10.1093/annonc/mdm140. [DOI] [PubMed] [Google Scholar]
- Haura EB, Tanvetyanon T, Chiappori A, Williams C, Simon G, Antonia S, et al. Phase I/II study of the Src inhibitor dasatinib in combination with erlotinib in advanced non-small-cell lung cancer. J Clin Oncol. 2010;28:1387–1394. doi: 10.1200/JCO.2009.25.4029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herbert C, Schieborr U, Saxena K, Juraszek J, De Smet F, Alcouffe C, et al. Molecular mechanism of SSR128129E, an extracellularly acting, small-molecule, allosteric inhibitor of FGF receptor signaling. Cancer Cell. 2013;23:489–501. doi: 10.1016/j.ccr.2013.02.018. [DOI] [PubMed] [Google Scholar]
- Hicklin DJ, Ellis LM. Role of the vascular endothelial growth factor pathway in tumor growth and angiogenesis. J Clin Oncol. 2005;23:1011–1027. doi: 10.1200/JCO.2005.06.081. [DOI] [PubMed] [Google Scholar]
- Hilberg F, Roth GJ, Krssak M, Kautschitsch S, Sommergruber W, Tontsch-Grunt U, et al. BIBF 1120: triple angiokinase inhibitor with sustained receptor blockade and good antitumor efficacy. Cancer Res. 2008;68:4774–4782. doi: 10.1158/0008-5472.CAN-07-6307. [DOI] [PubMed] [Google Scholar]
- Hillen F, Griffioen AW. Tumour vascularization: sprouting angiogenesis and beyond. Cancer Metastasis Rev. 2007;26:489–502. doi: 10.1007/s10555-007-9094-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Hinsbergh VW, Koolwijk P. Endothelial sprouting and angiogenesis: matrix metalloproteinases in the lead. Cardiovasc Res. 2008;78:203–212. doi: 10.1093/cvr/cvm102. [DOI] [PubMed] [Google Scholar]
- Holash J, Maisonpierre PC, Compton D, Boland P, Alexander CR, Zagzag D, et al. Vessel cooption, regression, and growth in tumors mediated by angiopoietins and VEGF. Science. 1999;284:1994–1998. doi: 10.1126/science.284.5422.1994. [DOI] [PubMed] [Google Scholar]
- Holden SN, Eckhardt SG, Basser R, de Boer R, Rischin D, Green M, et al. Clinical evaluation of ZD6474, an orally active inhibitor of VEGF and EGF receptor signaling, in patients with solid, malignant tumors. Ann Oncol. 2005;16:1391–1397. doi: 10.1093/annonc/mdi247. [DOI] [PubMed] [Google Scholar]
- Houck KA, Leung DW, Rowland AM, Winer J, Ferrara N. Dual regulation of vascular endothelial growth factor bioavailability by genetic and proteolytic mechanisms. J Biol Chem. 1992;267:26031–26037. [PubMed] [Google Scholar]
- Hsu C, Yang TS, Huo TI, Hsieh RK, Yu CW, Hwang WS, et al. Vandetanib in patients with inoperable hepatocellular carcinoma: a phase II, randomized, double-blind, placebo-controlled study. J Hepatol. 2012;56:1097–1103. doi: 10.1016/j.jhep.2011.12.013. [DOI] [PubMed] [Google Scholar]
- Hsu JY, Wakelee HA. Monoclonal antibodies targeting vascular endothelial growth factor: current status and future challenges in cancer therapy. BioDrugs. 2009;23:289–304. doi: 10.2165/11317600-000000000-00000. [DOI] [PubMed] [Google Scholar]
- Huang D, Ding Y, Li Y, Luo W-M, Zhang Z-F, Snider J, et al. Sunitinib acts primarily on tumor endothelium rather than tumor cells to inhibit the growth of renal cell carcinoma. Cancer Res. 2010;70:1053–1062. doi: 10.1158/0008-5472.CAN-09-3722. [DOI] [PubMed] [Google Scholar]
- Hurwitz H, Fehrenbacher L, Novotny W, Cartwright T, Hainsworth J, Heim W, et al. Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N Engl J Med. 2004;350:2335–2342. doi: 10.1056/NEJMoa032691. [DOI] [PubMed] [Google Scholar]
- Huveneers S, Truong H, Danen HJ. Integrins: signaling, disease, and therapy. Int J Radiat Biol. 2007;83:743–751. doi: 10.1080/09553000701481808. [DOI] [PubMed] [Google Scholar]
- Ijichi H, Chytil A, Gorska AE, Aakre ME, Bierie B, Tada M, et al. Inhibiting Cxcr2 disrupts tumor–stromal interactions and improves survival in a mouse model of pancreatic ductal adenocarcinoma. J Clin Invest. 2011;121:4106–4117. doi: 10.1172/JCI42754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivy SP, Wick JY, Kaufman BM. An overview of small-molecule inhibitors of VEGFR signaling. Nat Rev Clin Oncol. 2009;6:569–579. doi: 10.1038/nrclinonc.2009.130. [DOI] [PubMed] [Google Scholar]
- Izbicka E, Campos D, Carrizales G, Patnaik A. Biomarkers of anticancer activity of R115777 (Tipifarnib, Zarnestra) in human breast cancer models in vitro. Anticancer Res. 2005;25:3215–3223. [PubMed] [Google Scholar]
- Izzedine H, Ederhy S, Goldwasser F, Soria JC, Milano G, Cohen A, et al. Management of hypertension in angiogenesis inhibitor-treated patients. Ann Oncol. 2009;20:807–815. doi: 10.1093/annonc/mdn713. [DOI] [PubMed] [Google Scholar]
- Jain RK. Normalizing tumor vasculature with anti-angiogenic therapy: a new paradigm for combination therapy. Nat Med. 2001;7:987–989. doi: 10.1038/nm0901-987. [DOI] [PubMed] [Google Scholar]
- Jain RK. Molecular regulation of vessel maturation. Nat Med. 2003;9:685–693. doi: 10.1038/nm0603-685. [DOI] [PubMed] [Google Scholar]
- Jain RK. Normalization of tumor vasculature: an emerging concept in antiangiogenic therapy. Science. 2005;307:58–62. doi: 10.1126/science.1104819. [DOI] [PubMed] [Google Scholar]
- Jarvelainen H, Sainio A, Koulu M, Wight TN, Penttinen R. Extracellular matrix molecules: potential targets in pharmacotherapy. Pharmacol Rev. 2009;61:198–223. doi: 10.1124/pr.109.001289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang Y, Goldberg ID, Shi YE. Complex roles of tissue inhibitors of metalloproteinases in cancer. Oncogene. 2002;21:2245–2252. doi: 10.1038/sj.onc.1205291. [DOI] [PubMed] [Google Scholar]
- Johnson DH, Fehrenbacher L, Novotny WF, Herbst RS, Nemunaitis JJ, Jablons DM, et al. Randomized phase II trial comparing bevacizumab plus carboplatin and paclitaxel with carboplatin and paclitaxel alone in previously untreated locally advanced or metastatic non-small-cell lung cancer. J Clin Oncol. 2004;22:2184–2191. doi: 10.1200/JCO.2004.11.022. [DOI] [PubMed] [Google Scholar]
- Johnstone KD, Karoli T, Liu L, Dredge K, Copeman E, Li CP, et al. Synthesis and biological evaluation of polysulfated oligosaccharide glycosides as inhibitors of angiogenesis and tumor growth. J Med Chem. 2010;53:1686–1699. doi: 10.1021/jm901449m. [DOI] [PubMed] [Google Scholar]
- Joyce JA, Pollard JW. Microenvironmental regulation of metastasis. Nat Rev Cancer. 2009;9:239–252. doi: 10.1038/nrc2618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jubb AM, Oates AJ, Holden S, Koeppen H. Predicting benefit from anti-angiogenic agents in malignancy. Nat Rev Cancer. 2006;6:626–635. doi: 10.1038/nrc1946. [DOI] [PubMed] [Google Scholar]
- Kabbinavar F, Hurwitz HI, Fehrenbacher L, Meropol NJ, Novotny WF, Lieberman G, et al. Phase II, randomized trial comparing bevacizumab plus fluorouracil (FU)/leucovorin (LV) with FU/LV alone in patients with metastatic colorectal cancer. J Clin Oncol. 2003;21:60–65. doi: 10.1200/JCO.2003.10.066. [DOI] [PubMed] [Google Scholar]
- Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer. 2006;6:392–401. doi: 10.1038/nrc1877. [DOI] [PubMed] [Google Scholar]
- Karaca B, Kucukzeybek Y, Gorumlu G, Erten C, Gul MK, Cengiz E, et al. Profiling of angiogenic cytokines produced by hormone- and drug-refractory prostate cancer cell lines, PC-3 and DU-145 before and after treatment with gossypol. Eur Cytokine Netw. 2008;19:176–184. doi: 10.1684/ecn.2008.0139. [DOI] [PubMed] [Google Scholar]
- Karl E, Warner K, Zeitlin B, Kaneko T, Wurtzel L, Jin T, et al. Bcl-2 acts in a proangiogenic signaling pathway through nuclear factor-kappaB and CXC chemokines. Cancer Res. 2005;65:5063–5069. doi: 10.1158/0008-5472.CAN-05-0140. [DOI] [PubMed] [Google Scholar]
- Kerbel R, Folkman J. Clinical translation of angiogenesis inhibitors. Nat Rev Cancer. 2002;2:727–739. doi: 10.1038/nrc905. [DOI] [PubMed] [Google Scholar]
- Kerbel RS, Viloria-Petit A, Klement G, Rak J. ‘Accidental’ anti-angiogenic drugs. Anti-oncogene directed signal transduction inhibitors and conventional chemotherapeutic agents as examples. Eur J Cancer. 2000;36:1248–1257. doi: 10.1016/s0959-8049(00)00092-7. [DOI] [PubMed] [Google Scholar]
- Kim CK, Choi YK, Lee H, Ha KS, Won MH, Kwon YG, et al. The farnesyltransferase inhibitor LB42708 suppresses vascular endothelial growth factor-induced angiogenesis by inhibiting ras-dependent mitogen-activated protein kinase and phosphatidylinositol 3-kinase/Akt signal pathways. Mol Pharmacol. 2010;78:142–150. doi: 10.1124/mol.110.063586. [DOI] [PubMed] [Google Scholar]
- Kim R, Emi M, Arihiro K, Tanabe K, Uchida Y, Toge T. Chemosensitization by STI571 targeting the platelet-derived growth factor/platelet-derived growth factor receptor-signaling pathway in the tumor progression and angiogenesis of gastric carcinoma. Cancer. 2005;103:1800–1809. doi: 10.1002/cncr.20973. [DOI] [PubMed] [Google Scholar]
- Kimura Y, Okuda H. Resveratrol isolated from Polygonum cuspidatum root prevents tumor growth and metastasis to lung and tumor-induced neovascularization in Lewis lung carcinoma-bearing mice. J Nutr. 2001;131:1844–1849. doi: 10.1093/jn/131.6.1844. [DOI] [PubMed] [Google Scholar]
- Klenke FM, Abdollahi A, Bertl E, Gebhard MM, Ewerbeck V, Huber PE, et al. Tyrosine kinase inhibitor SU6668 represses chondrosarcoma growth via antiangiogenesis in vivo. BMC Cancer. 2007;7:49. doi: 10.1186/1471-2407-7-49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreisl TN, McNeill KA, Sul J, Iwamoto FM, Shih J, Fine HA. A phase I/II trial of vandetanib for patients with recurrent malignant glioma. Neuro Oncol. 2012;14:1519–1526. doi: 10.1093/neuonc/nos265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubota Y, Takubo K, Shimizu T, Ohno H, Kishi K, Shibuya M, et al. M-CSF inhibition selectively targets pathological angiogenesis and lymphangiogenesis. J Exp Med. 2009;206:1089–1102. doi: 10.1084/jem.20081605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lammerts E, Roswall P, Sundberg C, Gotwals PJ, Koteliansky VE, Reed RK, et al. Interference with TGF-beta1 and -beta3 in tumor stroma lowers tumor interstitial fluid pressure independently of growth in experimental carcinoma. Int J Cancer. 2002;102:453–462. doi: 10.1002/ijc.10722. [DOI] [PubMed] [Google Scholar]
- Lang SA, Klein D, Moser C, Gaumann A, Glockzin G, Dahlke MH, et al. Inhibition of heat shock protein 90 impairs epidermal growth factor-mediated signaling in gastric cancer cells and reduces tumor growth and vascularization in vivo. Mol Cancer Ther. 2007;6:1123–1132. doi: 10.1158/1535-7163.MCT-06-0628. [DOI] [PubMed] [Google Scholar]
- Leboulleux S, Bastholt L, Krause T, de la Fouchardiere C, Tennvall J, Awada A, et al. Vandetanib in locally advanced or metastatic differentiated thyroid cancer: a randomised, double-blind, phase 2 trial. Lancet Oncol. 2012;13:897–905. doi: 10.1016/S1470-2045(12)70335-2. [DOI] [PubMed] [Google Scholar]
- Ledermann JA, Hackshaw A, Kaye S, Jayson G, Gabra H, McNeish I, et al. Randomized phase II placebo-controlled trial of maintenance therapy using the oral triple angiokinase inhibitor BIBF 1120 after chemotherapy for relapsed ovarian cancer. J Clin Oncol. 2011;29:3798–3804. doi: 10.1200/JCO.2010.33.5208. [DOI] [PubMed] [Google Scholar]
- Lee J. Hypoxia-inducible Factor-1 (HIF-1)-independent hypoxia response of the small heat shock protein hsp-16.1 gene regulated by chromatin-remodeling factors in the nematode Caenorhabditis elegans. J Biol Chem. 2013;288:1582–1589. doi: 10.1074/jbc.M112.401554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JS, Hirsh V, Park K, Qin S, Blajman CR, Perng RP, et al. Vandetanib versus placebo in patients with advanced non-small-cell lung cancer after prior therapy with an epidermal growth factor receptor tyrosine kinase inhibitor: a randomized, double-blind phase III trial (ZEPHYR) J Clin Oncol. 2012;30:1114–1121. doi: 10.1200/JCO.2011.36.1709. [DOI] [PubMed] [Google Scholar]
- Leek RD, Talks KL, Pezzella F, Turley H, Campo L, Brown NS, et al. Relation of hypoxia-inducible factor-2 alpha (HIF-2 alpha) expression in tumor-infiltrative macrophages to tumor angiogenesis and the oxidative thymidine phosphorylase pathway in human breast cancer. Cancer Res. 2002;62:1326–1329. [PubMed] [Google Scholar]
- Leenders WP, Kusters B, de Waal RM. Vessel co-option: how tumors obtain blood supply in the absence of sprouting angiogenesis. Endothelium. 2002;9:83–87. doi: 10.1080/10623320212006. [DOI] [PubMed] [Google Scholar]
- Lewis CE, Pollard JW. Distinct role of macrophages in different tumor microenvironments. Cancer Res. 2006;66:605–612. doi: 10.1158/0008-5472.CAN-05-4005. [DOI] [PubMed] [Google Scholar]
- Li D, Williams JI, Pietras RJ. Squalamine and cisplatin block angiogenesis and growth of human ovarian cancer cells with or without HER-2 gene overexpression. Oncogene. 2002;21:2805–2814. doi: 10.1038/sj.onc.1205410. [DOI] [PubMed] [Google Scholar]
- Li WW, Li VW, Hutnik M, Chiou AS. Tumor angiogenesis as a target for dietary cancer prevention. J Oncol. 2012;2012:879623. doi: 10.1155/2012/879623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin EY, Li JF, Gnatovskiy L, Deng Y, Zhu L, Grzesik DA, et al. Macrophages regulate the angiogenic switch in a mouse model of breast cancer. Cancer Res. 2006;66:11238–11246. doi: 10.1158/0008-5472.CAN-06-1278. [DOI] [PubMed] [Google Scholar]
- Liu CJ, Lee PH, Lin DY, Wu CC, Jeng LB, Lin PW, et al. Heparanase inhibitor PI-88 as adjuvant therapy for hepatocellular carcinoma after curative resection: a randomized phase II trial for safety and optimal dosage. J Hepatol. 2009;50:958–968. doi: 10.1016/j.jhep.2008.12.023. [DOI] [PubMed] [Google Scholar]
- Llovet JM, Ricci S, Mazzaferro V, Hilgard P, Gane E, Blanc JF, et al. Sorafenib in advanced hepatocellular carcinoma. N Engl J Med. 2008;359:378–390. doi: 10.1056/NEJMoa0708857. [DOI] [PubMed] [Google Scholar]
- Loges S, Schmidt T, Carmeliet P. Mechanisms of resistance to anti-angiogenic therapy and development of third-generation anti-angiogenic drug candidates. Genes Cancer. 2010;1:12–25. doi: 10.1177/1947601909356574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez-Lopez C, LeRoith D, Torres-Aleman I. Insulin-like growth factor I is required for vessel remodeling in the adult brain. Proc Natl Acad Sci U S A. 2004;101:9833–9838. doi: 10.1073/pnas.0400337101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lord R, Nair S, Schache A, Spicer J, Somaihah N, Khoo V, et al. Low dose metronomic oral cyclophosphamide for hormone resistant prostate cancer: a phase II study. J Urol. 2007;177:2136–2140. doi: 10.1016/j.juro.2007.01.143. discussion 2140. [DOI] [PubMed] [Google Scholar]
- Lorusso G, Ruegg C. The tumor microenvironment and its contribution to tumor evolution toward metastasis. Histochem Cell Biol. 2008;130:1091–1103. doi: 10.1007/s00418-008-0530-8. [DOI] [PubMed] [Google Scholar]
- Ma J, Waxman DJ. Combination of antiangiogenesis with chemotherapy for more effective cancer treatment. Mol Cancer Ther. 2008;7:3670–3684. doi: 10.1158/1535-7163.MCT-08-0715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma J, Waxman DJ. Dominant effect of antiangiogenesis in combination therapy involving cyclophosphamide and axitinib. Clin Cancer Res. 2009;15:578–588. doi: 10.1158/1078-0432.CCR-08-1174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma J, Chen CS, Blute T, Waxman DJ. Antiangiogenesis enhances intratumoral drug retention. Cancer Res. 2011;71:2675–2685. doi: 10.1158/0008-5472.CAN-10-3242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacLauchlan S, Yu J, Parrish M, Asoulin TA, Schleicher M, Krady MM, et al. Endothelial nitric oxide synthase controls the expression of the angiogenesis inhibitor thrombospondin 2. Proc Natl Acad Sci U S A. 2011;108:E1137–E1145. doi: 10.1073/pnas.1104357108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeshima Y, Sudhakar A, Lively JC, Ueki K, Kharbanda S, Kahn CR, et al. Tumstatin, an endothelial cell-specific inhibitor of protein synthesis. Science. 2002;295:140–143. doi: 10.1126/science.1065298. [DOI] [PubMed] [Google Scholar]
- Manegold C, Vansteenkiste J, Cardenal F, Schuette W, Woll PJ, Ulsperger E, et al. Randomized phase II study of three doses of the integrin inhibitor cilengitide versus docetaxel as second-line treatment for patients with advanced non-small-cell lung cancer. Invest New Drugs. 2013;31:175–182. doi: 10.1007/s10637-012-9842-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manegold PC, Paringer C, Kulka U, Krimmel K, Eichhorn ME, Wilkowski R, et al. Antiangiogenic therapy with mammalian target of rapamycin inhibitor RAD001 (Everolimus) increases radiosensitivity in solid cancer. Clin Cancer Res. 2008;14:892–900. doi: 10.1158/1078-0432.CCR-07-0955. [DOI] [PubMed] [Google Scholar]
- Manthey CL, Johnson DL, Illig CR, Tuman RW, Zhou Z, Baker JF, et al. JNJ-28312141, a novel orally active colony-stimulating factor-1 receptor/FMS-related receptor tyrosine kinase-3 receptor tyrosine kinase inhibitor with potential utility in solid tumors, bone metastases, and acute myeloid leukemia. Mol Cancer Ther. 2009;8:3151–3161. doi: 10.1158/1535-7163.MCT-09-0255. [DOI] [PubMed] [Google Scholar]
- Martens E, Leyssen A, Van Aelst I, Fiten P, Piccard H, Hu J, et al. A monoclonal antibody inhibits gelatinase B/MMP-9 by selective binding to part of the catalytic domain and not to the fibronectin or zinc binding domains. Biochim Biophys Acta. 2007;1770:178–186. doi: 10.1016/j.bbagen.2006.10.012. [DOI] [PubMed] [Google Scholar]
- Matsuo Y, Sawai H, Ochi N, Yasuda A, Sakamoto M, Takahashi H, et al. Proteasome inhibitor MG132 inhibits angiogenesis in pancreatic cancer by blocking NF-kappaB activity. Dig Dis Sci. 2009;55:1167–1176. doi: 10.1007/s10620-009-0814-4. [DOI] [PubMed] [Google Scholar]
- May C, Doody JF, Abdullah R, Balderes P, Xu X, Chen CP, et al. Identification of a transiently exposed VE-cadherin epitope that allows for specific targeting of an antibody to the tumor neovasculature. Blood. 2005;105:4337–4344. doi: 10.1182/blood-2005-01-0010. [DOI] [PubMed] [Google Scholar]
- Meyerhardt JA, Ancukiewicz M, Abrams TA, Schrag D, Enzinger PC, Chan JA, et al. Phase I study of cetuximab, irinotecan, and vandetanib (ZD6474) as therapy for patients with previously treated metastastic colorectal cancer. PLoS One. 2012;7:e38231. doi: 10.1371/journal.pone.0038231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minor DR. Bevacizumab advanced melanoma (BEAM) was a positive trial. J Clin Oncol. 2012;30:2023. doi: 10.1200/JCO.2012.41.7477. author reply 2023–2024. [DOI] [PubMed] [Google Scholar]
- Mizukami Y, Kohgo Y, Chung DC. Hypoxia inducible factor-1 independent pathways in tumor angiogenesis. Clin Cancer Res. 2007;13:5670–5674. doi: 10.1158/1078-0432.CCR-07-0111. [DOI] [PubMed] [Google Scholar]
- Montero AJ, Escobar M, Lopes G, Gluck S, Vogel C. Bevacizumab in the treatment of metastatic breast cancer: friend or foe? Curr Oncol Rep. 2012;14:1–11. doi: 10.1007/s11912-011-0202-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moser C, Lang SA, Hackl C, Wagner C, Scheiffert E, Schlitt HJ, et al. Targeting HSP90 by the novel inhibitor NVP-AUY922 reduces growth and angiogenesis of pancreatic cancer. Anticancer Res. 2012;32:2551–2561. [PubMed] [Google Scholar]
- Mueller J, Gaertner FC, Blechert B, Janssen KP, Essler M. Targeting of tumor blood vessels: a phage-displayed tumor-homing peptide specifically binds to matrix metalloproteinase-2-processed collagen IV and blocks angiogenesis in vivo. Mol Cancer Res. 2009;7:1078–1085. doi: 10.1158/1541-7786.MCR-08-0538. [DOI] [PubMed] [Google Scholar]
- Mundel TM, Kalluri R. Type IV collagen-derived angiogenesis inhibitors. Microvasc Res. 2007;74:85–89. doi: 10.1016/j.mvr.2007.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murakami M. Lipid mediators in life science. Exp Anim. 2011;60:7–20. doi: 10.1538/expanim.60.7. [DOI] [PubMed] [Google Scholar]
- Murakami M, Nguyen LT, Hatanaka K, Schachterle W, Chen P-Y, Zhuang ZW, et al. FGF-dependent regulation of VEGF receptor 2 expression in mice. J Clin Invest. 2011;121:2668–2678. doi: 10.1172/JCI44762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murdoch C, Muthana M, Coffelt SB, Lewis CE. The role of myeloid cells in the promotion of tumour angiogenesis. Nat Rev Cancer. 2008;8:618–631. doi: 10.1038/nrc2444. [DOI] [PubMed] [Google Scholar]
- Murphy EA, Majeti BK, Barnes LA, Makale M, Weis SM, Lutu-Fuga K, et al. Nanoparticle-mediated drug delivery to tumor vasculature suppresses metastasis. Proc Natl Acad Sci U S A. 2008;105:9343–9348. doi: 10.1073/pnas.0803728105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagy JA, Feng D, Vasile E, Wong WH, Shih SC, Dvorak AM, et al. Permeability properties of tumor surrogate blood vessels induced by VEGF-A. Lab Invest. 2006;86:767–780. doi: 10.1038/labinvest.3700436. [DOI] [PubMed] [Google Scholar]
- Nakashima T, Huang C, Liu D, Kameyama K, Masuya D, Kobayashi S, et al. Neural-cadherin expression associated with angiogenesis in non-small-cell lung cancer patients. Br J Cancer. 2003;88:1727–1733. doi: 10.1038/sj.bjc.6600955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naumova E, Ubezio P, Garofalo A, Borsotti P, Cassis L, Riccardi E, et al. The vascular targeting property of paclitaxel is enhanced by SU6668, a receptor tyrosine kinase inhibitor, causing apoptosis of endothelial cells and inhibition of angiogenesis. Clin Cancer Res. 2006;12:1839–1849. doi: 10.1158/1078-0432.CCR-05-1615. [DOI] [PubMed] [Google Scholar]
- Navid F, Baker SD, McCarville MB, Stewart CF, Billups CA, Wu J, et al. Phase I and clinical pharmacology study of bevacizumab, sorafenib, and low-dose cyclophosphamide in children and young adults with refractory/recurrent solid tumors. Clin Cancer Res. 2013;19:236–246. doi: 10.1158/1078-0432.CCR-12-1897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nawrocki ST, Bruns CJ, Harbison MT, Bold RJ, Gotsch BS, Abbruzzese JL, et al. Effects of the proteasome inhibitor PS-341 on apoptosis and angiogenesis in orthotopic human pancreatic tumor xenografts. Mol Cancer Ther. 2002;1:1243–1253. [PubMed] [Google Scholar]
- Nelius T, Rinard K, Filleur S. Oral/metronomic cyclophosphamide-based chemotherapy as option for patients with castration–refractory prostate cancer: review of the literature. Cancer Treat Rev. 2011;37:444–455. doi: 10.1016/j.ctrv.2010.12.006. [DOI] [PubMed] [Google Scholar]
- Nyberg P, Xie L, Kalluri R. Endogenous inhibitors of angiogenesis. Cancer Res. 2005;65:3967–3979. doi: 10.1158/0008-5472.CAN-04-2427. [DOI] [PubMed] [Google Scholar]
- O'Reilly MS, Boehm T, Shing Y, Fukai N, Vasios G, Lane WS, et al. Endostatin: an endogenous inhibitor of angiogenesis and tumor growth. Cell. 1997;88:277–285. doi: 10.1016/s0092-8674(00)81848-6. [DOI] [PubMed] [Google Scholar]
- Ohtsu A, Shah MA, Van Cutsem E, Rha SY, Sawaki A, Park SR, et al. Bevacizumab in combination with chemotherapy as first-line therapy in advanced gastric cancer: a randomized, double-blind, placebo-controlled phase III study. J Clin Oncol. 2011;29:3968–3976. doi: 10.1200/JCO.2011.36.2236. [DOI] [PubMed] [Google Scholar]
- Pang X, Wu Y, Lu B, Chen J, Wang J, Yi Z, et al. (−)-Gossypol suppresses the growth of human prostate cancer xenografts via modulating VEGF signaling-mediated angiogenesis. Mol Cancer Ther. 2011;10:795–805. doi: 10.1158/1535-7163.MCT-10-0936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papageorgiou A, Kamat A, Benedict WF, Dinney C, McConkey DJ. Combination therapy with IFN-alpha plus bortezomib induces apoptosis and inhibits angiogenesis in human bladder cancer cells. Mol Cancer Ther. 2006;5:3032–3041. doi: 10.1158/1535-7163.MCT-05-0474. [DOI] [PubMed] [Google Scholar]
- Penel N, Adenis A, Bocci G. Cyclophosphamide-based metronomic chemotherapy: after 10 years of experience, where do we stand and where are we going? Crit Rev Oncol Hematol. 2012;82:40–50. doi: 10.1016/j.critrevonc.2011.04.009. [DOI] [PubMed] [Google Scholar]
- Polverini PJ. Angiogenesis in health and disease: insights into basic mechanisms and therapeutic opportunities. J Dent Educ. 2002;66:962–975. [PubMed] [Google Scholar]
- Presta M, Dell'Era P, Mitola S, Moroni E, Ronca R, Rusnati M. Fibroblast growth factor/fibroblast growth factor receptor system in angiogenesis. Cytokine Growth Factor Rev. 2005;16:159–178. doi: 10.1016/j.cytogfr.2005.01.004. [DOI] [PubMed] [Google Scholar]
- Pugh CW, Ratcliffe PJ. Regulation of angiogenesis by hypoxia: role of the HIF system. Nat Med. 2003;9:677–684. doi: 10.1038/nm0603-677. [DOI] [PubMed] [Google Scholar]
- Qian DZ, Wang X, Kachhap SK, Kato Y, Wei Y, Zhang L, et al. The histone deacetylase inhibitor NVP-LAQ824 inhibits angiogenesis and has a greater antitumor effect in combination with the vascular endothelial growth factor receptor tyrosine kinase inhibitor PTK787/ZK222584. Cancer Res. 2004;64:6626–6634. doi: 10.1158/0008-5472.CAN-04-0540. [DOI] [PubMed] [Google Scholar]
- Qin L, Zhao D, Xu J, Ren X, Terwilliger EF, Parangi S, et al. The vascular permeabilizing factors histamine and serotonin induce angiogenesis through TR3/Nur77 and subsequently truncate it through thrombospondin-1. Blood. 2013;121:2154–2164. doi: 10.1182/blood-2012-07-443903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reardon DA, Cheresh D. Cilengitide: a prototypic integrin inhibitor for the treatment of glioblastoma and other malignancies. Genes Cancer. 2011;2:1159–1165. doi: 10.1177/1947601912450586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ribatti D. Endogenous inhibitors of angiogenesis: a historical review. Leuk Res. 2009;33:638–644. doi: 10.1016/j.leukres.2008.11.019. [DOI] [PubMed] [Google Scholar]
- Ribatti D, Djonov V. Intussusceptive microvascular growth in tumors. Cancer Lett. 2012;316:126–131. doi: 10.1016/j.canlet.2011.10.040. [DOI] [PubMed] [Google Scholar]
- Rogers TL, Holen I. Tumour macrophages as potential targets of bisphosphonates. J Transl Med. 2011;9:177. doi: 10.1186/1479-5876-9-177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roskoski R., Jr Sunitinib: a VEGF and PDGF receptor protein kinase and angiogenesis inhibitor. Biochem Biophys Res Commun. 2007;356:323–328. doi: 10.1016/j.bbrc.2007.02.156. [DOI] [PubMed] [Google Scholar]
- Rothwell PM, Wilson M, Elwin CE, Norrving B, Algra A, Warlow CP, et al. Long-term effect of aspirin on colorectal cancer incidence and mortality: 20-year follow-up of five randomised trials. Lancet. 2010;376:1741–1750. doi: 10.1016/S0140-6736(10)61543-7. [DOI] [PubMed] [Google Scholar]
- Rudge JS, Holash J, Hylton D, Russell M, Jiang S, Leidich R, et al. VEGF Trap complex formation measures production rates of VEGF, providing a biomarker for predicting efficacious angiogenic blockade. Proc Natl Acad Sci U S A. 2007;104:18363–18370. doi: 10.1073/pnas.0708865104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruoslahti E, Bhatia SN, Sailor MJ. Targeting of drugs and nanoparticles to tumors. J Cell Biol. 2010;188:759–768. doi: 10.1083/jcb.200910104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sabbadini RA. Sphingosine-1-phosphate antibodies as potential agents in the treatment of cancer and age-related macular degeneration. Br J Pharmacol. 2011;162:1225–1238. doi: 10.1111/j.1476-5381.2010.01118.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanderson S, Valenti M, Gowan S, Patterson L, Ahmad Z, Workman P, et al. Benzoquinone ansamycin heat shock protein 90 inhibitors modulate multiple functions required for tumor angiogenesis. Mol Cancer Ther. 2006;5:522–532. doi: 10.1158/1535-7163.MCT-05-0439. [DOI] [PubMed] [Google Scholar]
- Santini D, Vespasiani Gentilucci U, Vincenzi B, Picardi A, Vasaturo F, La Cesa A, et al. The antineoplastic role of bisphosphonates: from basic research to clinical evidence. Ann Oncol. 2003;14:1468–1476. doi: 10.1093/annonc/mdg401. [DOI] [PubMed] [Google Scholar]
- Santini D, Vincenzi B, Galluzzo S, Battistoni F, Rocci L, Venditti O, et al. Repeated intermittent low-dose therapy with zoledronic acid induces an early, sustained, and long-lasting decrease of peripheral vascular endothelial growth factor levels in cancer patients. Clin Cancer Res. 2007;13(15 Pt 1):4482–4486. doi: 10.1158/1078-0432.CCR-07-0551. [DOI] [PubMed] [Google Scholar]
- Schuster C, Eikesdal HP, Puntervoll H, Geisler J, Geisler S, Heinrich D, et al. Clinical efficacy and safety of bevacizumab monotherapy in patients with metastatic melanoma: predictive importance of induced early hypertension. PLoS One. 2012;7:e38364. doi: 10.1371/journal.pone.0038364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz EL. Antivascular actions of microtubule-binding drugs. Clin Cancer Res. 2009;15:2594–2601. doi: 10.1158/1078-0432.CCR-08-2710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahrzad S, Shirasawa S, Sasazuki T, Rak JW, Coomber BL. Low-dose metronomic cyclophosphamide treatment mediates ischemia-dependent K-ras mutation in colorectal carcinoma xenografts. Oncogene. 2008;27:3729–3738. doi: 10.1038/sj.onc.1211031. [DOI] [PubMed] [Google Scholar]
- Shaked Y, Ciarrocchi A, Franco M, Lee CR, Man S, Cheung AM, et al. Therapy-induced acute recruitment of circulating endothelial progenitor cells to tumors. Science. 2006;313:1785–1787. doi: 10.1126/science.1127592. [DOI] [PubMed] [Google Scholar]
- Shinohara ET, Cao C, Niermann K, Mu Y, Zeng F, Hallahan DE, et al. Enhanced radiation damage of tumor vasculature by mTOR inhibitors. Oncogene. 2005;24:5414–5422. doi: 10.1038/sj.onc.1208715. [DOI] [PubMed] [Google Scholar]
- Shojaei F, Wu X, Zhong C, Yu L, Liang XH, Yao J, et al. Bv8 regulates myeloid-cell-dependent tumour angiogenesis. Nature. 2007;450:825–831. doi: 10.1038/nature06348. [DOI] [PubMed] [Google Scholar]
- Shojaei F, Singh M, Thompson JD, Ferrara N. Role of Bv8 in neutrophil-dependent angiogenesis in a transgenic model of cancer progression. Proc Natl Acad Sci U S A. 2008;105:2640–2645. doi: 10.1073/pnas.0712185105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stacchiotti S, Negri T, Zaffaroni N, Palassini E, Morosi C, Brich S, et al. Sunitinib in advanced alveolar soft part sarcoma: evidence of a direct antitumor effect. Ann Oncol. 2011;22:1682–1690. doi: 10.1093/annonc/mdq644. [DOI] [PubMed] [Google Scholar]
- Stathopoulos GT, Moschos C, Loutrari H, Kollintza A, Psallidas I, Karabela S, et al. Zoledronic acid is effective against experimental malignant pleural effusion. Am J Respir Crit Care Med. 2008;178:50–59. doi: 10.1164/rccm.200710-1513OC. [DOI] [PubMed] [Google Scholar]
- Strieter RM, Belperio JA, Phillips RJ, Keane MP. CXC chemokines in angiogenesis of cancer. Semin Cancer Biol. 2004;14:195–200. doi: 10.1016/j.semcancer.2003.10.006. [DOI] [PubMed] [Google Scholar]
- Summy JM, Trevino JG, Lesslie DP, Baker CH, Shakespeare WC, Wang Y, et al. AP23846, a novel and highly potent Src family kinase inhibitor, reduces vascular endothelial growth factor and interleukin-8 expression in human solid tumor cell lines and abrogates downstream angiogenic processes. Mol Cancer Ther. 2005;4:1900–1911. doi: 10.1158/1535-7163.MCT-05-0171. [DOI] [PubMed] [Google Scholar]
- Sunwoo JB, Chen Z, Dong G, Yeh N, Crowl Bancroft C, Sausville E, et al. Novel proteasome inhibitor PS-341 inhibits activation of nuclear factor-kappa B, cell survival, tumor growth, and angiogenesis in squamous cell carcinoma. Clin Cancer Res. 2001;7:1419–1428. [PubMed] [Google Scholar]
- Taeger J, Moser C, Hellerbrand C, Mycielska ME, Glockzin G, Schlitt HJ, et al. Targeting FGFR/PDGFR/VEGFR impairs tumor growth, angiogenesis, and metastasis by effects on tumor cells, endothelial cells, and pericytes in pancreatic cancer. Mol Cancer Ther. 2011;10:2157–2167. doi: 10.1158/1535-7163.MCT-11-0312. [DOI] [PubMed] [Google Scholar]
- Takahashi H, Shibuya M. The vascular endothelial growth factor (VEGF)/VEGF receptor system and its role under physiological and pathological conditions. Clin Sci. 2005;109:227–241. doi: 10.1042/CS20040370. [DOI] [PubMed] [Google Scholar]
- Tang Y, Nakada MT, Kesavan P, McCabe F, Millar H, Rafferty P, et al. Extracellular matrix metalloproteinase inducer stimulates tumor angiogenesis by elevating vascular endothelial cell growth factor and matrix metalloproteinases. Cancer Res. 2005;65:3193–3199. doi: 10.1158/0008-5472.CAN-04-3605. [DOI] [PubMed] [Google Scholar]
- Tei K, Kawakami-Kimura N, Taguchi O, Kumamoto K, Higashiyama S, Taniguchi N, et al. Roles of cell adhesion molecules in tumor angiogenesis induced by cotransplantation of cancer and endothelial cells to nude rats. Cancer Res. 2002;62:6289–6296. [PubMed] [Google Scholar]
- Teicher BA. A systems approach to cancer therapy. (Antioncogenics + standard cytotoxics-->mechanism(s) of interaction) Cancer Metastasis Rev. 1996;15:247–272. doi: 10.1007/BF00437479. [DOI] [PubMed] [Google Scholar]
- di Tomaso E, London N, Fuja D, Logie J, Tyrrell JA, Kamoun W, et al. PDGF-C induces maturation of blood vessels in a model of glioblastoma and attenuates the response to anti-VEGF treatment. PLoS One. 2009;4:e5123. doi: 10.1371/journal.pone.0005123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ton GN, Banaszynski ME, Kolesar JM. Vandetanib: a novel targeted therapy for the treatment of metastatic or locally advanced medullary thyroid cancer. Am J Health Syst Pharm. 2013;70:849–855. doi: 10.2146/ajhp120253. [DOI] [PubMed] [Google Scholar]
- Tong RT, Boucher Y, Kozin SV, Winkler F, Hicklin DJ, Jain RK. Vascular normalization by vascular endothelial growth factor receptor 2 blockade induces a pressure gradient across the vasculature and improves drug penetration in tumors. Cancer Res. 2004;64:3731–3736. doi: 10.1158/0008-5472.CAN-04-0074. [DOI] [PubMed] [Google Scholar]
- Trepel J, Mollapour M, Giaccone G, Neckers L. Targeting the dynamic HSP90 complex in cancer. Nat Rev Cancer. 2010;10:537–549. doi: 10.1038/nrc2887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsujii M, Kawano S, Tsuji S, Sawaoka H, Hori M, DuBois RN. Cyclooxygenase regulates angiogenesis induced by colon cancer cells. Cell. 1998;93:705–716. doi: 10.1016/s0092-8674(00)81433-6. [DOI] [PubMed] [Google Scholar]
- Van Cutsem E, de Haas S, Kang YK, Ohtsu A, Tebbutt NC, Ming Xu J, et al. Bevacizumab in combination with chemotherapy as first-line therapy in advanced gastric cancer: a biomarker evaluation from the AVAGAST randomized phase III trial. J Clin Oncol. 2012;30:2119–2127. doi: 10.1200/JCO.2011.39.9824. [DOI] [PubMed] [Google Scholar]
- Veltman JD, Lambers ME, van Nimwegen M, Hendriks RW, Hoogsteden HC, Hegmans JP, et al. Zoledronic acid impairs myeloid differentiation to tumour-associated macrophages in mesothelioma. Br J Cancer. 2010;103:629–641. doi: 10.1038/sj.bjc.6605814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vergote IB, Colombo N, Kutarska E, Del Campo J, Pippitt C, Casado A, et al. Phase II study comparing volociximab (an antiangiogenic antibody) and pegylated liposomal doxorubicin (PLD) with PLD alone in recurrent ovarian or primary peritoneal cancer. ASCO Meeting Abstracts. 2009;27((15S)):5560. [Google Scholar]
- Verheul HM, Pinedo HM. Possible molecular mechanisms involved in the toxicity of angiogenesis inhibition. Nat Rev Cancer. 2007;7:475–485. doi: 10.1038/nrc2152. [DOI] [PubMed] [Google Scholar]
- Vermorken JB, Guigay J, Mesia R, Trigo JM, Keilholz U, Kerber A, et al. Phase I/II trial of cilengitide with cetuximab, cisplatin and 5-fluorouracil in recurrent and/or metastatic squamous cell cancer of the head and neck: findings of the phase I part. Br J Cancer. 2012;104:1691–1696. doi: 10.1038/bjc.2011.152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visentin B, Vekich JA, Sibbald BJ, Cavalli AL, Moreno KM, Matteo RG, et al. Validation of an anti-sphingosine-1-phosphate antibody as a potential therapeutic in reducing growth, invasion, and angiogenesis in multiple tumor lineages. Cancer Cell. 2006;9:225–238. doi: 10.1016/j.ccr.2006.02.023. [DOI] [PubMed] [Google Scholar]
- Vlodavsky I, Friedmann Y. Molecular properties and involvement of heparanase in cancer metastasis and angiogenesis. J Clin Invest. 2001;108:341–347. doi: 10.1172/JCI13662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang D, Dubois RN. Eicosanoids and cancer. Nat Rev Cancer. 2010;10:181–193. doi: 10.1038/nrc2809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang D, Wang H, Brown J, Daikoku T, Ning W, Shi Q, et al. CXCL1 induced by prostaglandin E2 promotes angiogenesis in colorectal cancer. J Exp Med. 2006;203:941–951. doi: 10.1084/jem.20052124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L, Park H, Chhim S, Ding Y, Jiang W, Queen C, et al. A novel monoclonal antibody to fibroblast growth factor 2 effectively inhibits growth of hepatocellular carcinoma xenografts. Mol Cancer Ther. 2012;11:864–872. doi: 10.1158/1535-7163.MCT-11-0813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z, Song W, Aboukameel A, Mohammad M, Wang G, Banerjee S, et al. TW-37, a small-molecule inhibitor of Bcl-2, inhibits cell growth and invasion in pancreatic cancer. Int J Cancer. 2008;123:958–966. doi: 10.1002/ijc.23610. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Wang Z, Chui WK, Ho PC. Nanoparticulate delivery system targeted to tumor neovasculature for combined anticancer and antiangiogenesis therapy. Pharm Res. 2011;28:585–596. doi: 10.1007/s11095-010-0308-2. [DOI] [PubMed] [Google Scholar]
- Wedge SR, Ogilvie DJ, Dukes M, Kendrew J, Chester R, Jackson JA, et al. ZD6474 inhibits vascular endothelial growth factor signaling, angiogenesis, and tumor growth following oral administration. Cancer Res. 2002;62:4645–4655. [PubMed] [Google Scholar]
- Wei D, Wang L, He Y, Xiong HQ, Abbruzzese JL, Xie K. Celecoxib inhibits vascular endothelial growth factor expression in and reduces angiogenesis and metastasis of human pancreatic cancer via suppression of Sp1 transcription factor activity. Cancer Res. 2004;64:2030–2038. doi: 10.1158/0008-5472.can-03-1945. [DOI] [PubMed] [Google Scholar]
- Weis SM, Cheresh DA. Alphav integrins in angiogenesis and cancer. Cold Spring Harb Perspect Med. 2011;1:a006478. doi: 10.1101/cshperspect.a006478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wells SA, Jr, Robinson BG, Gagel RF, Dralle H, Fagin JA, Santoro M, et al. Vandetanib in patients with locally advanced or metastatic medullary thyroid cancer: a randomized, double-blind phase III trial. J Clin Oncol. 2012;30:134–141. doi: 10.1200/JCO.2011.35.5040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitelock JM, Murdoch AD, Iozzo RV, Underwood PA. The degradation of human endothelial cell-derived perlecan and release of bound basic fibroblast growth factor by stromelysin, collagenase, plasmin, and heparanases. J Biol Chem. 1996;271:10079–10086. doi: 10.1074/jbc.271.17.10079. [DOI] [PubMed] [Google Scholar]
- Willis LM, El-Remessy AB, Somanath PR, Deremer DL, Fagan SC. Angiotensin receptor blockers and angiogenesis: clinical and experimental evidence. Clin Sci. 2011;120:307–319. doi: 10.1042/cs20100389. [DOI] [PubMed] [Google Scholar]
- Yao M, Zhou W, Sangha S, Albert A, Chang AJ, Liu TC, et al. Effects of nonselective cyclooxygenase inhibition with low-dose ibuprofen on tumor growth, angiogenesis, metastasis, and survival in a mouse model of colorectal cancer. Clin Cancer Res. 2005;11:1618–1628. doi: 10.1158/1078-0432.CCR-04-1696. [DOI] [PubMed] [Google Scholar]
- Yoshikawa D, Ojima H, Kokubu A, Ochiya T, Kasai S, Hirohashi S, et al. Vandetanib (ZD6474), an inhibitor of VEGFR and EGFR signalling, as a novel molecular-targeted therapy against cholangiocarcinoma. Br J Cancer. 2009;100:1257–1266. doi: 10.1038/sj.bjc.6604988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang CC, Yan Z, Zhang Q, Kuszpit K, Zasadny K, Qiu M, et al. PF-03732010: a fully human monoclonal antibody against P-cadherin with antitumor and antimetastatic activity. Clin Cancer Res. 2010;16:5177–5188. doi: 10.1158/1078-0432.CCR-10-1343. [DOI] [PubMed] [Google Scholar]
- Zhao G, Li W-Y, Chen D, Henry JR, Li H-Y, Chen Z, et al. A novel, selective inhibitor of fibroblast growth factor receptors that shows a potent broad spectrum of antitumor activity in several tumor xenograft models. Mol Cancer Ther. 2011;10:2200–2210. doi: 10.1158/1535-7163.MCT-11-0306. [DOI] [PubMed] [Google Scholar]
- Zhao WM, Wang L, Park H, Chhim S, Tanphanich M, Yashiro M, et al. Monoclonal antibodies to fibroblast growth factor receptor 2 effectively inhibit growth of gastric tumor xenografts. Clin Cancer Res. 2010;16:5750–5758. doi: 10.1158/1078-0432.CCR-10-0531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou H, Roy S, Cochran E, Zouaoui R, Chu CL, Duffner J, et al. M402, a novel heparan sulfate mimetic, targets multiple pathways implicated in tumor progression and metastasis. PLoS One. 2011;6:e21106. doi: 10.1371/journal.pone.0021106. [DOI] [PMC free article] [PubMed] [Google Scholar]