

Abstract
All cells have the capacity to accumulate neutral lipids and package them into lipid droplets. Recent proteomic analyses indicate that lipid droplets are not simple lipid storage depots, but rather complex organelles that have multiple cellular functions. One of these proposed functions is to distribute neutral lipids as well as phospholipids to various membrane-bound organelles within the cell. Here, we summarize the lipid droplet-associated membrane-trafficking proteins and review the evidence that lipid droplets interact with endoplasmic reticulum, endosomes, peroxisomes, and mitochondria. Based on this evidence, we present a model for how lipid droplets can distribute lipids to specific membrane compartments.
Keywords: Cell organelle, Lipid droplets, Traffic
1 Introduction
Dysregulation in neutral lipid storage has been linked to a variety of metabolic diseases. Excessive storage is associated with obesity, type II diabetes, hepatic steatosis, and atherosclerosis while lack of neutral lipid storage is associated with lipodystrophies. Understanding the molecular mechanisms that regulate neutral lipid storage holds the key to developing therapeutic treatments for these metabolic diseases.
Virtually all bacterial and eukaryotic cells are able to accumulate neutral lipids and store them in lipid droplets that generally have a spherical appearance, contain various species of neutral lipids and are surrounded by a monolayer of phospholipid with embedded integral as well as peripheral proteins [1, 2]. Similar structures are also found to be secreted by some cells in the form of lipoproteins and milk fat globules. Lipid droplets were among the earliest intracellular structures observed using microscope. As a result, morphologists have assigned many names to these structures, some of which portray a misleading if not inaccurate picture of their structure and function. This morphologic discord is made worse by the fact that lipoproteins also belong to this class of cellular structures [1]. Now there appears to be some degree of agreement that cytoplasmic lipid droplets are organelles and not static inclusions [2]. We have argued there needs to be a unifying hypothesis that embraces all of these names and, at the same time, focuses research on the fundamental issue in lipid storage: how are these containers synthesized and what is the range of their cellular function [3]. For this reason, we presented for consideration the adiposome hypothesis. This hypothesis proposes that cells contain cellular machinery (the adiposome) that produces cytoplasmic lipid storage containers (adiposome droplets) as well as lipid export containers (adiposome lipoproteins). Thus, adiposome droplets are lipid-storing organelles that move around in cells on microtubule tracks and distribute lipids to various membrane systems using specific docking and undocking machinery. However, as most researchers in the field prefer the term lipid droplet, we will conform to this practice for the remainder of the review. Outlined below is some of the evidence in the literature that supports the hypothesis that lipid droplets transport lipids to other cellular organelles by the interaction between lipid droplets and these organelles. Other aspects of lipid droplet biology, including biogenesis, lipid biosynthesis and catabolism, signal transduction, energy and cholesterol homeostasis and protein storage/degradation have previously been reviewed [4-14].
2 Lipid droplet proteome and genome are rich in membrane traffic proteins
Several different families of abundant lipid droplet proteins have been identified during the past 20 years. The best studied are the PAT [Perilipin, adipocyte differentiation related protein (ADRP) or Adipophilin, tail-interacting protein of 47 kDa (TIP47)] family of proteins found in mammals [9] and insects [10] and the oleosins found in plants [12]. The availability of these marker proteins has been critical for developing protocols to purify lipid droplets from yeast, plant, insect, and mammalian cells, and further analyze them using proteomics [3, 15-28]. These studies resulted in a variety of proteins that regulate membrane traffic (Table 1). There are five groups of these proteins: (i) small GTPases that regulate vesicle formation, motility, targeting, and tethering; (ii) motor proteins such as kinesin and myosin that move vesicles on the cytoskeleton; (iii) soluble NSF attachment receptor (SNARE) [α-soluble NSF attachment protein (SNAP) and SNAP29]) and N-ethylmaleimide-sensitive factor (NSF) that drive membrane docking and fusion; (iv) vesicular traffic proteins (Arf1, membrin, Sar1, Sec21, COPs, and Sec22) that regulate cargo sorting and vesicle budding; and (v) membrane traffic proteins of unknown function (VAT1, Rap1, P22, and TIP47). In several studies, enrichment of this class of proteins was further documented by immunoblotting [3, 27]. The presence of membrane-traffic proteins in lipid droplets supports the view that they are dynamic organelles specialized for bi-directional, inter-membrane lipid transport.
Table 1.
Lipid droplet-associated membrane traffic proteins identified by proteomic analysesa)
| CHOK2-1[3] | CHOK2-2[27] | 3T3L1[18] | A431[19] | U937[26] | HuH7[17] | Liver[20] | HepG2[21] | Drob)[24] | Yeast[23] | |
|---|---|---|---|---|---|---|---|---|---|---|
| Rab1 | + | + | + | + | + | + | + | |||
| Rab2 | + | + | ||||||||
| Rab3 | + | |||||||||
| Rab3 GEF | + | |||||||||
| Rab5 | + | + | + | + | + | + | + | |||
| Rab6 | + | |||||||||
| Rab7 | + | + | + | + | + | + | + | + | ||
| Rab8 | + | + | + | |||||||
| Rab10 | + | + | + | + | + | |||||
| Rab11 | + | + | + | + | + | |||||
| Rab14 | + | + | + | + | ||||||
| Rab18 | + | + | + | + | + | + | ||||
| Rab19 | + | |||||||||
| Rab21 | + | |||||||||
| Rab24 | + | |||||||||
| Rab33 | + | |||||||||
| Rab34 | + | |||||||||
| Rab35 | + | + | ||||||||
| Rab39 | + | |||||||||
| Rab41 | + | |||||||||
| Sec21 | + | |||||||||
| Sec22 | + | + | ||||||||
| α-SNAP | + | |||||||||
| SNAP29 | + | |||||||||
| Arf1 | + | + | ||||||||
| ArfGAP1 | + | |||||||||
| ARL10C | + | |||||||||
| Membrin | + | |||||||||
| αCop | + | |||||||||
| βCop | + | |||||||||
| γCop | + | |||||||||
| PLD | + | |||||||||
| Sar1 | + | + | + | + | ||||||
| VAT-1 | + | + | + | |||||||
| Rap1 | + | + | ||||||||
| TIP47 | + | + | + | + | + | |||||
| P22 | + | + | + | |||||||
| Myosin | + | + | ||||||||
| Kinesin | + | |||||||||
| Trans- | + | |||||||||
| membrane | ||||||||||
| Trafficking | ||||||||||
| protein | ||||||||||
| NSF | + | |||||||||
| Vesicle | + | |||||||||
| Transport- | ||||||||||
| related | ||||||||||
| Protein | ||||||||||
| VPS20 | + | |||||||||
| VPS26 | + | |||||||||
| RER1 Protein | + | |||||||||
| β-Adaptin | + | |||||||||
| Clathrin Heavy | + | |||||||||
| Chain |
Note: The proteins in Supporting Information of these papers are not included.
Drosophila.
The interpretation of proteomic data can be complicated by contaminating proteins that accumulate during purification. Furthermore, proteins may have distinct functions on different organelles, which can be misleading. One approach to addressing these problems is to compare all published lipid droplet proteomes and see if any of the proteins of interest show up in more than one study. Table 1 lists the membrane traffic proteins identified in lipid droplet proteomes from yeast, Drosophila and mammals. Among the mammal proteomes, nearly all the studies found various isoforms of Rab GTPase (e.g. eight studies found Rab7 and seven found Rab5), VAT1 and TIP47. More impressive, perhaps, is that proteins like Sar1 and Rab1 were found in lipid droplet proteomes from all three groups of cells. The fact that the same proteins often appear in multiple studies across species supports the view that membrane-traffic proteins are functionally associated with lipid droplets. This conclusion is further supported by yeast genetics. Szymanski et al. [29] carried out a detailed screen for genes that were essential for normal lipid droplet morphology in Saccharomyces cerevisiae. They identified 58 essential genes, 16 of which were genes coding for proteins involved in endosomal-lysosomal traffic. A recent functional genomic screen also showed that Arf1 and its coatomer COPI regulate lipid droplet formation [30].
3 Cytoskeleton-dependent lipid droplet traffic
Lipid droplets are capable of rapid, microtubule-dependent movement as shown with live-cell imaging of the Drosophila embryo [31] and mammalian HuH-7 cells [32]. As with vesicles, the movement is driven by the plus-end and minus-end microtubule motor proteins, kinesin and dynein [33]. The localization of these motor proteins on lipid droplets was determined both by co-immunoprecipitation (IP) with the lipid droplet protein ADRP [34] and by immunofluorescence [33]. Furthermore, Erk-mediated phosphorylation of dynein [35] increases the association of this motor protein with lipid droplets as does the phosphatase inhibitor, vanadate. Interestingly, the ability of lipid droplets to form and grow in size is dependent on motor proteins and microtubules. Dynein neutralizing antibodies reduce lipid droplet formation [35] and depolymerization of microtubules with nocodazole inhibits homotypic fusion of lipid droplets. Lipid droplets may also engage other cytoskeleton systems since recent proteomic studies identified additional cytoskeleton proteins such as actin and plectin on purified droplets as well as the motor protein myosin [20, 27]. Importantly, GTP regulates the association of cytoskeleton proteins with lipid droplets (see Section 4).
4 Targeting of lipid droplets to membrane-bound organelles
The ability of lipid droplets to move bi-directional on microtubules must be important because genetic disruption of lipid storage droplet (LSD)2, a Drosophila PAT family member protein that regulates microtubule motor proteins [36], reduces lipid accumulation [37]. Most likely lipid droplet movement is part of a system that delivers lipid droplets to various membrane organelles in the cell for lipid exchange. The evidence that lipid droplets interact with various organelles is extensive and compelling.
4.1 Endoplasmic reticulum
An interaction between lipid droplets and ER has been observed by electron microscope (EM) in studies from yeast to mammals [29, 38, 39]. Some might disregard the ER as a lipid droplet target because there is considerable evidence that the lipid droplet biogenesis machinery resides in this organelle. Numerous studies suggest, however, there are regulated interactions between lipid droplets and ER that are independent of lipid droplet formation. One of the clearest morphologic examples of ER-lipid droplet interactions is in macrophages loaded with cholesterol and cholesterol esters [40]. Embedded in these lipid droplets are whorls of smooth ER that are continuous with elements of rough ER. Moreover, the ER whorls appear to be the location of esterases that de-esterify cholesterol esters, thereby creating a pool of free cholesterol in the lipid droplet. Perhaps more compelling evidence of lipid droplet-ER interaction is the studies showing that the GTP/GDP state of Rab18 regulates the interaction between the two organelles [41, 42]. Finally, hepatitis C virus appears to replicate in regions of ER that are juxtaposed to lipid droplets [43]. These observations suggest that regions of lipid droplet-ER interaction create a microenvironment favorable for virus replication. Perhaps this environment is needed for bi-directional movement of lipids between the two organelles during virus replication.
4.2 Endosomes
Several lines of research suggest that a functional interaction between lipid droplets and endosomes is an important aspect of normal membrane traffic. Electron microscopy has found that the endosomes of cells taking up high-density lipoprotein (HDL) form a close association with lipid droplets [44]. Moreover, immunofluorescence detects an association between the HDL receptor (SRB1) and lipid droplets after HDL internalization [45]. This interaction is consistent with the fact that the lipid droplet proteome contains a number of proteins involved in mediating and regulating endosomal membrane traffic (Table 1). In addition, immunogold labeling has localized Rab5 and Rab11 on the surface of isolated lipid droplets [46]. This suggests that Rab GTPases regulate the interaction between lipid droplets and endosomes. Indeed, purified lipid droplets are able to recruit the Rab5 effector, early endosome antigen 1 (EEA1), in vitro, and when Rab proteins are removed from both purified lipid droplets and early endosomes using RabGDI, lipid droplets are no longer able to tightly associate with purified endosomes [46]. These results provide direct evidence that Rab5 as well as other members of the Rab family regulate lipid droplet-endosome interactions.
Lipid droplets also contain proteins known to mediate SNARE-dependent membrane-membrane fusion (Table 1). Indeed, Bostrom et al. [47] recently reported that SNAREs mediate homotypic fusion between droplets. By contrast, lipid droplet-endosome interaction in vitro does not appear to result in fusion between the two organelles. To the contrary, ATP causes dissociation of bound endosomes from isolated lipid droplets [46], suggesting that lipid droplets have the ability to dock but not fuse with endosomes. Another membrane traffic regulator on lipid droplets is the GTPase Arf1. It both recruits phospholipase D (PLD) [48, 49] and regulates ADRP binding. GTP Arf1 facilitates ADRP binding while GDP-bound Arf1 dissociates ADRP from lipid droplets [50]. Lipid droplets are also able to recruit Arf1 in vitro, as well as coatomers and EEA1 [27, 46].
4.3 Peroxisomes
One of the earliest observations of an interaction between lipid droplets and peroxisomes was in EM images of 3T3 L1 adipocytes by Novikoff [38]. EM studies have also documented an interaction between lipid droplets and peroxisomes following stimulation of steroidogenic luteal cells with luteinizing hormone (LH) [51]. Live-cell imaging has confirmed that the interaction between peroxisomes and lipid droplets is extremely dynamic [52]. The relative lack of quantification as well as the low resolution of these studies, however, cannot rule out that these observations were fortuitous and not representative of the in situ behavior of these organelles. Nevertheless, in a recent study using the yeast Saccharomyces cerevisiae Binns et al. [23] provide clear evidence that peroxisomes interact with lipid droplets. A mutant yeast strain with peroxisomes that are unable to oxidize fatty acids was found to make intimate contact with lipid droplets. In some cases, peroxisomes were found in contact with lipid droplets that had formed processes called “gnarls” that extended into the core of the lipid droplet. Thus, in wild type cells lipid droplet-peroxisome contacts may facilitate the coupling of lipolysis on the lipid droplet to fatty acid oxidation in the peroxisome [23].
4.4 Mitochondria
Several lipid droplet proteomic studies found proteins typically associated with the mitochondria. While we cannot rule out the possibility that these are contaminants, interaction between these two organelles is expected because in mammalian cells fatty acid oxidation occurs in mitochondria. In fact, an association between lipid droplets and mitochondria has been reported in EM studies of adipocytes [38, 53], liver cells [54], lactating cells from mice and rats [39]. In developing porcine oocytes fluorescence resonance energy transfer found that mitochondria and lipid droplets are within 6-10 nm of each other [55]. Lipid droplets and mitochondria colocalize in skeletal muscle cells [56] where most lipids are utilized for energy production and this interaction is enhanced when energy requirements are increased [57]. Finally, deletion studies suggest that caveolae protein caveolin-1 plays a role in maintaining the interaction between lipid droplets and mitochondria [58].
Although the molecular mechanisms governing the interactions between lipid droplets and various cellular organelles are not known, the evidence for these interactions is compelling. Future studies need to focus on identifying the molecular machinery that mediates these interactions, the regulatory machinery that controls these processes, and the various external and internal cues that promote these interactions.
4.5 Lipid droplet-mediated intracellular lipid traffic
The reported instances of lipid droplets interacting with various cellular organelles are numerous and widespread, but the nature and purpose of these interactions remain unclear. We have argued that one function is inter-membrane lipid delivery and processing [46]. The lipid composition of lipid droplets is complex. In Chinese hamster ovary (CHO) K2 cells, for example, the lipid droplet contains more than 160 species of phospholipid (including lyso and ether-linked species), 100 species of neutral lipid as well as special lipids such as monoalk(en)yl diacylglycerol (MADAG) [59]. This diversity implies lipid droplets play a central role in managing membrane lipids. They also provide clues about which lipids are linked to these interactions. For example, the lipid composition of CHO cell lipid droplet suggests a role in ether lipid metabolism. The first three enzymes in ether lipid biosynthesis are in peroxisomes and lipid droplets in cells defective in peroxisome biogenesis do not contain MADAG [59]. This implies that MADAG precursors are transferred to lipid droplets through a direct interaction with peroxisome. Most likely, the 1-alkyl-2-acyl-glycerol is converted to MADAG (possibly by a 1,2-diacylglycerol acyltransferase) and then stored in lipid droplets. Lipid droplets may function as a supply depot for ether phospholipid biosynthesis as well as a recovery organelle for storage, much like the inter-conversion of phospholipids and triglyceride. The significant amount of lysophospholipids in lipid droplets [59, 60] also indicates a role in phospholipid recycling.
We have proposed that each Rab species on lipid droplets regulates interaction with a specific membrane system [46]. For example, Rab5 controls interaction with early endosomes and Rab18 with ER. Mitochondria and peroxisomes may use Rab species specific for these two compartments. In support of this hypothesis, recent proteomic studies have determined that both purified synaptic vesicles [61] and purified secretory membranes [62] contain multiple Rab species. We think these interactions necessarily lead to the formation of transient inter-compartmental contact sites (TICCS) that transfer lipids, small molecules and ions without membrane fusion (reviewed in [46]). TICCS may be involved in the interaction of ER subdomains with mitochondria and the transfer of phosphatidylserine (PtdSer) for conversion to phosphatidylethanolamine (PtdEtn) [63]. A similar interaction occurs between ER and plasma membrane that appears to be important for lipid transfer [64, 65]. TICCS may also mediate interaction between lipid droplets and peroxisomes. Recent studies suggest how TICCS might work (Fig. 1). Sterols can move from the cell surface (Fig. 1, yellow line) through endosomal (Fig. 1, blue circle) membranes to lipid droplets (Fig. 1, yellow circle) by an ATP-independent pathway [66]. The binding of early endosomes to lipid droplets also does not require ATP [46], which raises the possibility that a series of TICCS, which can connect together multiple endosomal compartments through recognition sites (Fig. 1, red squares), allows the passage of sterols from the cell surface to the lipid droplet without energy expenditure. Such a mechanism could mediate bi-directional, regulated movement of cholesterol between HDL (and other lipoproteins) in endosomes and internal compartments.
Figure 1.
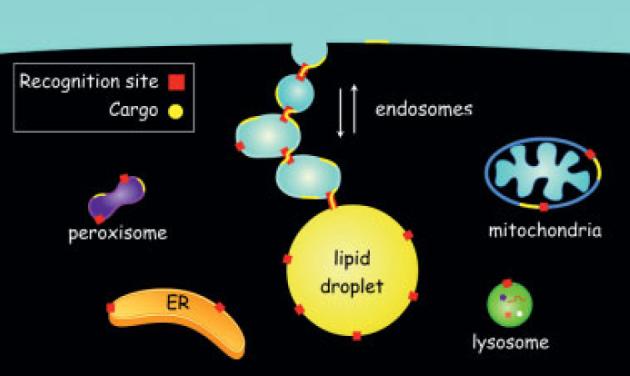
Transient Inter-Compartmental Contact Sites (TICCS) mediate lipid traffic between various membrane compartments and lipid droplets. The figure represents TICCS hypothesis. The top blue area is extracellular compartment. Cargo lipids (yellow line for lipids on membrane and yellow circle for lipids in lipid droplets) move between the cell surface and lipid droplets using endosomal intermediates (blue circular compartment). Migration is bi-directional and depends on the formation endosome-endosome as well as endosome-lipid droplet TICCS. Recognition sites (red square) for each organelle contain the molecular machinery necessary for the formation of TICCS. Compartmental specificity of TICCS formation is regulated by Rab-GTP (from [46] with permission).
Identifying the molecular basis of TICCS formation is an important future goal in understanding how lipid droplets interact with other organelles. Except mediating homotypic fusion between lipid droplets, the presence of SNARE proteins and NSF in purified lipid droplets suggests these proteins may have docking activity to drive the formation of TICCS. Arf1 and its coatamers may also play a novel role in regulating certain aspect of TICCS that are involved in lipid droplet formation. Of course, it is possible that multiple mechanisms are involved in lipid droplet-mediated intracellular lipid trafficking. Lipid droplet-associated SNARE proteins may also drive fusion between lipid droplets and certain cellular organelles.
5 Concluding remarks
Recent studies have revealed that lipid droplets, previously considered static storage depots of cellular neutral lipids, are organelles actively engaged in lipid metabolism, lipid storage, membrane traffic, protein degradation and cellular signaling. Lipid transportation may be the central function of this organelle by delivering and acquiring lipids via TICCS. Knowledge and tools are being developed that will enable us to learn more about the function of this organelle in health and disease. This new perspective offers a fresh new understanding of how to develop cellular strategies for treating lipid storage diseases.
Acknowledgments
Authors would like to thank Drs. Peter Michaely and Dorothy Mundy for very useful discussions and suggestions and Angela Diehl for the artwork. This work is supported by grants from the National Basic Research Program of China (Grants 2004CB720000 and 2006CB911001), National Institutes of Health (HL 20948, GM 52016, GM 70117), the Perot Family Foundation and the Cecil H. Green Distinguished Chair in Cellular and Molecular Biology.
Abbreviations
- ADRP
adipocyte differentiation related protein
- CHO
Chinese hamster ovary
- EEA1
early endosome antigen 1
- EM
electron microscope
- HDL
high-density lipoprotein
- NSF
N-ethylmaleimide-sensitive factor
- PAT
Perilipin, ADRP or Adipophilin, TIP47
- SNAP
soluble NSF attachment protein
- SNARE
soluble NSF attachment receptor
- TICCS
transient inter-compartmental contact sites
- TIP47
tail-interacting protein of 47 kDa
Footnotes
The authors have declared no conflict of intrest.
6 References
- [1].Murphy DJ. The biogenesis and functions of lipid bodies in animals, plants and microorganisms. Prog. Lipid Res. 2001;40:325–438. doi: 10.1016/s0163-7827(01)00013-3. [DOI] [PubMed] [Google Scholar]
- [2].Martin S, Parton RG. Lipid droplets: a unified view of a dynamic organelle. Nat. Rev. Mol. Cell. Biol. 2006;7:373–378. doi: 10.1038/nrm1912. [DOI] [PubMed] [Google Scholar]
- [3].Liu P, Ying Y, Zhao Y, Mundy DI, et al. Chinese hamster ovary K2 cell lipid droplets appear to be metabolic organelles involved in membrane traffic. J. Biol. Chem. 2004;279:3787–3792. doi: 10.1074/jbc.M311945200. [DOI] [PubMed] [Google Scholar]
- [4].Zweytick D, Athenstaedt K, Daum G. Intracellular lipid particles of eukaryotic cells. Biochim. Biophys. Acta. 2000;1469:101–120. doi: 10.1016/s0005-2736(00)00294-7. [DOI] [PubMed] [Google Scholar]
- [5].Brown DA. Lipid droplets: proteins floating on a pool of fat. Curr. Biol. 2001;11:R446–R449. doi: 10.1016/s0960-9822(01)00257-3. [DOI] [PubMed] [Google Scholar]
- [6].Waltermann M, Steinbuchel A. Neutral lipid bodies in prokaryotes: recent insights into structure, formation, and relationship to eukaryotic lipid depots. J. Bacteriol. 2005;187:3607–3619. doi: 10.1128/JB.187.11.3607-3619.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Wolins NE, Brasaemle DL, Bickel PE. A proposed model of fat packaging by exchangeable lipid droplet proteins. FEBS Lett. 2006;580:5484–5491. doi: 10.1016/j.febslet.2006.08.040. [DOI] [PubMed] [Google Scholar]
- [8].Greenberg AS, Obin MS. Many roads lead to the lipid droplet. Cell Metab. 2008;7:472–473. doi: 10.1016/j.cmet.2008.05.005. [DOI] [PubMed] [Google Scholar]
- [9].Londos C, Brasaemle DL, Schultz CJ, Segrest JP, Kimmel AR. Perilipins, ADRP, and other proteins that associate with intracellular neutral lipid droplets in animal cells. Semin. Cell Dev. Biol. 1999;10:51–58. doi: 10.1006/scdb.1998.0275. [DOI] [PubMed] [Google Scholar]
- [10].Lu X, Gruia-Gray J, Copeland NG, Gilbert DJ, et al. The murine perilipin gene: the lipid droplet-associated perilipins derive from tissue-specific, mRNA splice variants and define a gene family of ancient origin. Mamm. Genome. 2001;12:741–749. doi: 10.1007/s00335-01-2055-5. [DOI] [PubMed] [Google Scholar]
- [11].Ducharme NA, Bickel PE. Lipid droplets in lipogenesis and lipolysis. Endocrinology. 2008;149:942–949. doi: 10.1210/en.2007-1713. [DOI] [PubMed] [Google Scholar]
- [12].Murphy DJ, Keen JN, O'Sullivan JN, Au DM, et al. A class of amphipathic proteins associated with lipid storage bodies in plants. Possible similarities with animal serum apolipoproteins. Biochim. Biophys. Acta. 1991;1088:86–94. doi: 10.1016/0167-4781(91)90156-g. [DOI] [PubMed] [Google Scholar]
- [13].Melo RC, Sabban A, Weller PF. Leukocyte lipid bodies: inflammation-related organelles are rapidly detected by wet scanning electron microscopy. J. Lipid Res. 2006;47:2589–2594. doi: 10.1194/jlr.D600028-JLR200. [DOI] [PubMed] [Google Scholar]
- [14].Fujimoto T, Ohsaki Y, Cheng J, Suzuki M, Shinohara Y. Lipid droplets: a classic organelle with new outfits. Histochem. Cell Biol. 2008;130:263–279. doi: 10.1007/s00418-008-0449-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Athenstaedt K, Zweytick D, Jandrositz A, Kohlwein SD, Daum G. Identification and characterization of major lipid particle proteins of the yeast Saccharomyces cerevisiae. J. Bacteriol. 1999;181:6441–6448. doi: 10.1128/jb.181.20.6441-6448.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Wu CC, Howell KE, Neville MC, Yates JR, 3rd, McManaman JL. Proteomics reveal a link between the endoplasmic reticulum and lipid secretory mechanisms in mammary epithelial cells. Electrophoresis. 2000;21:3470–3482. doi: 10.1002/1522-2683(20001001)21:16<3470::AID-ELPS3470>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- [17].Fujimoto Y, Itabe H, Sakai J, Makita M, et al. Identification of major proteins in the lipid droplet-enriched fraction isolated from the human hepatocyte cell line HuH7. Biochim. Biophys. Acta. 2004;1644:47–59. doi: 10.1016/j.bbamcr.2003.10.018. [DOI] [PubMed] [Google Scholar]
- [18].Brasaemle DL, Dolios G, Shapiro L, Wang R. Proteomic analysis of proteins associated with lipid droplets of basal and lipolytically stimulated 3T3-L1 adipocytes. J. Biol. Chem. 2004;279:46835–46842. doi: 10.1074/jbc.M409340200. [DOI] [PubMed] [Google Scholar]
- [19].Umlauf E, Csaszar E, Moertelmaier M, Schuetz GJ, et al. Association of stomatin with lipid bodies. J. Biol. Chem. 2004;279:23699–23709. doi: 10.1074/jbc.M310546200. [DOI] [PubMed] [Google Scholar]
- [20].Turro S, Ingelmo-Torres M, Estanyol JM, Tebar F, et al. Identification and characterization of associated with lipid droplet protein 1: A novel membrane-associated protein that resides on hepatic lipid droplets. Traffic. 2006;7:1254–1269. doi: 10.1111/j.1600-0854.2006.00465.x. [DOI] [PubMed] [Google Scholar]
- [21].Sato S, Fukasawa M, Yamakawa Y, Natsume T, et al. Proteomic profiling of lipid droplet proteins in hepatoma cell lines expressing hepatitis C virus core protein. J. Biochem. (Tokyo) 2006;139:921–930. doi: 10.1093/jb/mvj104. [DOI] [PubMed] [Google Scholar]
- [22].Cermelli S, Guo Y, Gross SP, Welte MA. The lipid-droplet proteome reveals that droplets are a protein-storage depot. Curr. Biol. 2006;16:1783–1795. doi: 10.1016/j.cub.2006.07.062. [DOI] [PubMed] [Google Scholar]
- [23].Binns D, Januszewski T, Chen Y, Hill J, et al. An intimate collaboration between peroxisomes and lipid bodies. J. Cell Biol. 2006;173:719–731. doi: 10.1083/jcb.200511125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Beller M, Riedel D, Jansch L, Dieterich G, et al. Characterization of the Drosophila lipid droplet subproteome. Mol. Cell. Proteomics. 2006;5:1082–1094. doi: 10.1074/mcp.M600011-MCP200. [DOI] [PubMed] [Google Scholar]
- [25].Kim SC, Chen Y, Mirza S, Xu Y, et al. A clean, more efficient method for in-solution digestion of protein mixtures without detergent or urea. J. Proteome Res. 2006;5:3446–3452. doi: 10.1021/pr0603396. [DOI] [PubMed] [Google Scholar]
- [26].Wan HC, Melo RC, Jin Z, Dvorak AM, Weller PF. Roles and origins of leukocyte lipid bodies: proteomic and ultrastructural studies. FASEB J. 2007;21:167–178. doi: 10.1096/fj.06-6711com. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Bartz R, Zehmer JK, Zhu M, Chen Y, et al. Dynamic activity of lipid droplets: protein phosphorylation and GTP-mediated protein translocation. J. Proteome Res. 2007;6:3256–3265. doi: 10.1021/pr070158j. [DOI] [PubMed] [Google Scholar]
- [28].Katavic V, Agrawal GK, Hajduch M, Harris SL, Thelen JJ. Protein and lipid composition analysis of oil bodies from two Brassica napus cultivars. Proteomics. 2006;6:4586–4598. doi: 10.1002/pmic.200600020. [DOI] [PubMed] [Google Scholar]
- [29].Szymanski KM, Binns D, Bartz R, Grishin NV, et al. The lipodystrophy protein seipin is found at endoplasmic reticulum lipid droplet junctions and is important for droplet morphology. Proc. Natl. Acad. Sci. USA. 2007;104:20890–20895. doi: 10.1073/pnas.0704154104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Guo Y, Walther TC, Rao M, Stuurman N, et al. Functional genomic screen reveals genes involved in lipid-droplet formation and utilization. Nature. 2008;453:657–661. doi: 10.1038/nature06928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Welte MA, Gross SP, Postner M, Block SM, Wieschaus EF. Developmental regulation of vesicle transport in Drosophila embryos: forces and kinetics. Cell. 1998;92:547–557. doi: 10.1016/s0092-8674(00)80947-2. [DOI] [PubMed] [Google Scholar]
- [32].Targett-Adams P, Chambers D, Gledhill S, Hope RG, et al. Live cell analysis and targeting of the lipid droplet-binding adipocyte differentiation-related protein. J. Biol. Chem. 2003;278:15998–16007. doi: 10.1074/jbc.M211289200. [DOI] [PubMed] [Google Scholar]
- [33].Gross SP, Welte MA, Block SM, Wieschaus EF. Dynein-mediated cargo transport in vivo. A switch controls travel distance. J. Cell Biol. 2000;148:945–956. doi: 10.1083/jcb.148.5.945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Bostrom P, Rutberg M, Ericsson J, Holmdahl P, et al. Cytosolic lipid droplets increase in size by microtubule-dependent complex formation. Arterioscler. Thromb. Vasc. Biol. 2005;25:1945–1951. doi: 10.1161/01.ATV.0000179676.41064.d4. [DOI] [PubMed] [Google Scholar]
- [35].Andersson L, Bostrom P, Ericson J, Rutberg M, et al. PLD1 and ERK2 regulate cytosolic lipid droplet formation. J. Cell Sci. 2006;119:2246–2257. doi: 10.1242/jcs.02941. [DOI] [PubMed] [Google Scholar]
- [36].Welte MA, Cermelli S, Griner J, Viera A, et al. Regulation of lipid-droplet transport by the perilipin homolog LSD2. Curr. Biol. 2005;15:1266–1275. doi: 10.1016/j.cub.2005.06.062. [DOI] [PubMed] [Google Scholar]
- [37].Teixeira L, Rabouille C, Rorth P, Ephrussi A, Vanzo NF. Drosophila Perilipin/ADRP homologue Lsd2 regulates lipid metabolism. Mech. Dev. 2003;120:1071–1081. doi: 10.1016/s0925-4773(03)00158-8. [DOI] [PubMed] [Google Scholar]
- [38].Novikoff AB, Novikoff PM, Rosen OM, Rubin CS. Organelle relationships in cultured 3T3-L1 preadipocytes. J. Cell Biol. 1980;87:180–196. doi: 10.1083/jcb.87.1.180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Stemberger BH, Walsh RM, Patton S. Morphometric evaluation of lipid droplet associations with secretory vesicles, mitochondria and other components in the lactating cell. Cell Tissue Res. 1984;236:471–475. doi: 10.1007/BF00214252. [DOI] [PubMed] [Google Scholar]
- [40].McGookey DJ, Anderson RG. Morphological characterization of the cholesteryl ester cycle in cultured mouse macrophage foam cells. J. Cell Biol. 1983;97:1156–1168. doi: 10.1083/jcb.97.4.1156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Ozeki S, Cheng J, Tauchi-Sato K, Hatano N, et al. Rab18 localizes to lipid droplets and induces their close apposition to the endoplasmic reticulum-derived membrane. J. Cell Sci. 2005;118:2601–2611. doi: 10.1242/jcs.02401. [DOI] [PubMed] [Google Scholar]
- [42].Martin S, Driessen K, Nixon SJ, Zerial M, Parton RG. Regulated localization of Rab18 to lipid droplets: effects of lipolytic stimulation and inhibition of lipid droplet catabolism. J. Biol. Chem. 2005;280:42325–42335. doi: 10.1074/jbc.M506651200. [DOI] [PubMed] [Google Scholar]
- [43].Targett-Adams P, Boulant S, McLauchlan J. Visualization of double-stranded RNA in cells supporting hepatitis C virus RNA replication. J. Virol. 2008;82:2182–2195. doi: 10.1128/JVI.01565-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Schmitz G, Robenek H, Lohmann U, Assmann G. Interaction of high density lipoproteins with cholesteryl esterladen macrophages: biochemical and morphological characterization of cell surface receptor binding, endocytosis and resecretion of high density lipoproteins by macrophages. EMBO J. 1985;4:613–622. doi: 10.1002/j.1460-2075.1985.tb03674.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Frolov A, Petrescu A, Atshaves BP, So PT, et al. High density lipoprotein-mediated cholesterol uptake and targeting to lipid droplets in intact L-cell fibroblasts. A single- and multiphoton fluorescence approach. J. Biol. Chem. 2000;275:12769–12780. doi: 10.1074/jbc.275.17.12769. [DOI] [PubMed] [Google Scholar]
- [46].Liu P, Bartz R, Zehmer JK, Ying YS, et al. Rab-regulated interaction of early endosomes with lipid droplets. Biochim. Biophys. Acta. 2007;1773:784–793. doi: 10.1016/j.bbamcr.2007.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Bostrom P, Andersson L, Rutberg M, Perman J, et al. SNARE proteins mediate fusion between cytosolic lipid droplets and are implicated in insulin sensitivity. Nat. Cell Biol. 2007;9:1286–1293. doi: 10.1038/ncb1648. [DOI] [PubMed] [Google Scholar]
- [48].Marchesan D, Rutberg M, Andersson L, Asp L, et al. A phospholipase D-dependent process forms lipid droplets containing caveolin, adipocyte differentiation-related protein, and vimentin in a cell-free system. J. Biol. Chem. 2003;278:27293–27300. doi: 10.1074/jbc.M301430200. [DOI] [PubMed] [Google Scholar]
- [49].Nakamura N, Banno Y, Tamiya-Koizumi K. Arf1-dependent PLD1 is localized to oleic acid-induced lipid droplets in NIH3T3 cells. Biochem. Biophys. Res. Commun. 2005;335:117–123. doi: 10.1016/j.bbrc.2005.07.050. [DOI] [PubMed] [Google Scholar]
- [50].Nakamura N, Akashi T, Taneda T, Kogo H, et al. ADRP is dissociated from lipid droplets by ARF1-dependent mechanism. Biochem. Biophys. Res. Commun. 2004;322:957–965. doi: 10.1016/j.bbrc.2004.08.010. [DOI] [PubMed] [Google Scholar]
- [51].Mendis-Handagama SM. Peroxisomes and intracellular cholesterol trafficking in adult rat Leydig cells following lLuteinizing hormone stimulation. Tissue Cell. 2000;32:102–106. doi: 10.1054/tice.1999.0092. [DOI] [PubMed] [Google Scholar]
- [52].Schrader M. Tubulo-reticular clusters of peroxisomes in living COS-7 cells: dynamic behavior and association with lipid droplets. J. Histochem. Cytochem. 2001;49:1421–1429. doi: 10.1177/002215540104901110. [DOI] [PubMed] [Google Scholar]
- [53].Blanchette-Mackie EJ, Scow RO. Movement of lipolytic products to mitochondria in brown adipose tissue of young rats: an electron microscope study. J. Lipid Res. 1983;24:229–244. [PubMed] [Google Scholar]
- [54].Fadeeva M. M. K. a. E. O. Ultrastructural study of liver cells from rooks living in ecologically unfavorable areas. Biology Bulletin. 2006;33:99–106. [PubMed] [Google Scholar]
- [55].Sturmey RG, O'Toole PJ, Leese HJ. Fluorescence resonance energy transfer analysis of mitochondrial:lipid association in the porcine oocyte. Reproduction. 2006;132:829–837. doi: 10.1530/REP-06-0073. [DOI] [PubMed] [Google Scholar]
- [56].Shaw CS, Jones DA, Wagenmakers AJ. Network distribution of mitochondria and lipid droplets in human muscle fibres. Histochem. Cell Biol. 2008;129:65–72. doi: 10.1007/s00418-007-0349-8. [DOI] [PubMed] [Google Scholar]
- [57].Tarnopolsky MA, Rennie CD, Robertshaw HA, Fedak-Tarnopolsky SN, et al. Influence of endurance exercise training and sex on intramyocellular lipid and mitochondrial ultrastructure, substrate use, and mitochondrial enzyme activity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007;292:R1271–1278. doi: 10.1152/ajpregu.00472.2006. [DOI] [PubMed] [Google Scholar]
- [58].Cohen AW, Razani B, Schubert W, Williams TM, et al. Role of caveolin-1 in the modulation of lipolysis and lipid droplet formation. Diabetes. 2004;53:1261–1270. doi: 10.2337/diabetes.53.5.1261. [DOI] [PubMed] [Google Scholar]
- [59].Bartz R, Li WH, Venables B, Zehmer JK, et al. Lipidomics reveals that adiposomes store ether lipids and mediate phospholipid traffic. J. Lipid Res. 2007;48:837–847. doi: 10.1194/jlr.M600413-JLR200. [DOI] [PubMed] [Google Scholar]
- [60].Tauchi-Sato K, Ozeki S, Houjou T, Taguchi R, Fujimoto T. The surface of lipid droplets is a phospholipid monolayer with a unique fatty acid composition. J. Biol. Chem. 2002;277:44507–44512. doi: 10.1074/jbc.M207712200. [DOI] [PubMed] [Google Scholar]
- [61].Takamori S, Holt M, Stenius K, Lemke EA, et al. Molecular anatomy of a trafficking organelle. Cell. 2006;127:831–846. doi: 10.1016/j.cell.2006.10.030. [DOI] [PubMed] [Google Scholar]
- [62].Gilchrist A, Au CE, Hiding J, Bell AW, et al. Quantitative proteomics analysis of the secretory pathway. Cell. 2006;127:1265–1281. doi: 10.1016/j.cell.2006.10.036. [DOI] [PubMed] [Google Scholar]
- [63].Voelker DR. Interorganelle transport of aminoglycerophospholipids. Biochim. Biophys. Acta. 2000;1486:97–107. doi: 10.1016/s1388-1981(00)00051-2. [DOI] [PubMed] [Google Scholar]
- [64].Pichler H, Gaigg B, Hrastnik C, Achleitner G, et al. A subfraction of the yeast endoplasmic reticulum associates with the plasma membrane and has a high capacity to synthesize lipids. Eur. J. Biochem. 2001;268:2351–2361. doi: 10.1046/j.1432-1327.2001.02116.x. [DOI] [PubMed] [Google Scholar]
- [65].Schnabl M, Daum G, Pichler H. Multiple lipid transport pathways to the plasma membrane in yeast. Biochim. Biophys. Acta. 2005;1687:130–140. doi: 10.1016/j.bbalip.2004.11.016. [DOI] [PubMed] [Google Scholar]
- [66].Wustner D, Mondal M, Tabas I, Maxfield FR. Direct observation of rapid internalization and intracellular transport of sterol by macrophage foam cells. Traffic. 2005;6:396–412. doi: 10.1111/j.1600-0854.2005.00285.x. [DOI] [PubMed] [Google Scholar]