

Abstract
Microarray technology provides a unique opportunity to examine gene expression patterns in human embryonic stem cells (hESCs). We performed a meta-analysis of 38 original studies reporting on the transcriptome of hESCs. We determined that 1076 genes were found overexpressed in hESCs by at least 3 studies when compared to differentiated cell types, thus composing a “consensus hESC gene list”. Only one gene was reported by all studies: the homeodomain transcription factor POU5F1/OCT3/4. The list comprised other genes critical for pluripotency such as the transcription factors NANOG and SOX2, and the growth factors TDGF1/CRIPTO and Galanin. We show that CD24 and SEMA6A, two cell surface protein-coding genes from the top of the consensus hESC gene list, display a strong and specific membrane protein expression on hESCs. Moreover, CD24 labeling permits to purify by flow cytometry hESCs co-cultured on human fibroblasts. The “consensus hESC gene list” also included the FZD7 WNT receptor, the G protein-coupled receptor GPR19, and the HELLS helicase which could play an important role in hESCs biology. Conversely, we identified 783 genes downregulated in hESCs and reported in at least three studies. This “consensus differentiation gene list” included the IL6ST/GP130 LIF receptor. We created an online hESC expression atlas, (http://amazonia.montp.inserm.fr), to provide an easy access to this public transcriptome dataset. Expression histograms comparing hESC to a broad collection of fetal and adult tissues can be retrieved with this web tool for more than 15 000 genes.
Keywords: pluripotent stem cells, gene expression profiling, microarray analysis
Introduction
In the preimplantation mammalian embryo, the inner cell mass is able to differentiate into any cell type of the embryo proper. It has been recognized in mice since 1981 that embryonic stem cells (ESCs) with a prolonged proliferative capacity in vitro can be derived from the inner cell mass [1]. ESC line derivation from human embryos was reported in 1998 [2]. ESCs are pluripotent cells that can contribute to all tissues in vivo, and to the three primary germ layers as well as extraembryonic tissues in vitro. As pluripotency is maintained even after prolonged periods of culture, human ESCs (hESCs) have a great therapeutic potential in regenerative medicine. Careful molecular characterization of this unique cellular model of pluripotency should help to optimize and scale up in vitro manipulation of hESCs for clinical applications. Some genes specific to the very early stages of development and expressed in hESC lines such as POU5F1/OCT3/4, NANOG, REX1, SOX2, FGF4 and FOXD3 have already been identified [3–8]. However, the picture is far from complete and the molecular mechanisms involved in self-renewal and pluripotency are still under tight scrutiny [9, 10]. Moreover, extensive knowledge on hESCs should help in designing protocols for the isolation and production of other multi/pluripotent stem cells derived from tissues of adult individuals.
Microarrays are a major technical breakthrough that can monitor the expression of a whole genome in one experiment. Application of this technology to hESCs has largely contributed to our knowledge on the mechanisms underlying the maintenance of pluripotency of hESCs and their in vitro differentiation. Unfortunately, the datasets generated are heterogeneous both in accessibility (public databases or Supplemental data) and in the techniques used (variability in microarray design, sample labeling techniques, choice of control samples and computational tools). Despite our findings of 38 original publications reporting hESC transcriptome analyses, the disparities between datasets, the variety of sources, the substantial know-how needed for transcriptome data mining discourage most non-specialists to consult this information. Thus, large amount of data are very much underused, and lack alternative interpretations since only the conclusions reflecting the analysis carried out by the authors are presented. This situation has led to initiatives such as ONCOMINE in the field of cancer [11]. We present here the first effort in compiling all publicly available microarray data relating to hESCs. From the 38 original publications studying the hESC transcriptome, we identified genes that were consistently overexpressed in hESCs when compared to differentiated samples (“consensus hESC gene list”) and underexpressed in hESCs (“consensus differentiation gene list”) in different studies. These lists will further deepen our knowledge on this unique cell model of developmental biology. Concurrently, we created an on-line database, Amazonia!, which provides an easy access to this public transcriptome dataset.
Materials and Methods
Lists of genes differentially expressed
Analyzing 38 original studies using transcriptome analysis to study hESC, we were able to collect 20 lists of transcripts that were upregulated in hESCs compared to differentiated cell types and 11 lists of transcripts that were downregulated (see Supplemental Tables S1, S2 and S3). We only selected transcripts lists that provided a fold ratio of the mean expression in hESCs to that in differentiated cells. Each list was mapped to Unigene build 176. When the mean value of expression in differentiated cells was zero, which occurred with ESTs and SAGE, the hESC/differentiation ratio was arbitrarily set at 50. Only genes with a fold ratio greater or equal to 2 (“hESC” genes), or lower or equal to 0.5 (“differentiation” genes) were selected. The Gene Ontology annotation analysis was carried out using the Fatigo+ tool on the Babelomics website (http://babelomics.bioinfo.cipf.es) using gene symbols [12]. Only annotation with a false discovery rates (FDR) adjusted P-value < 0.05 were considered significant.
Integrating Affymetrix GeneChip datasets obtained from distinct studies
In order to compare the transcriptome of hESCs to that of differentiated cell populations, we built an expression compendium by combining the U133A (Affymetrix, Santa Clara, USA) microarray data from 8 publications [13–20]. Indeed, we and others have shown that the GeneChip system (Affymetrix) allows direct comparison between datasets obtained in different centers, provided that the same chip and the same normalization are used [10, 21, 22]. The number of samples amounted to 217, including 24 hESC samples (11 different hESC lines). All samples were normalized before analysis with the GCOS 1.2 software (Affymetrix), using the “global scaling” method with a TGT value set to 100. The “ detection call” can either be “present”, when the perfect match probes are significantly more hybridized than the mismatch probes, “absent” when both perfect match and mismatch probes display a similar fluorescent signal, or “marginal” when the probeset does neither comply to the present nor to the absent call criteria. When several probesets measured the same gene, only the probeset with the maximal number of present detection call across all samples was selected. This step reduced the list of probesets to 14 074. This dataset is available as Supplemental Table S4 and can be accessed on our website http://amazonia.montp.inserm.fr.
Hierarchical clustering
Hierarchical clustering was carried out on the detection call data with the CLUSTER and TREEVIEW software packages [23]. The value 1 was assigned to present calls, −1 to absent calls and 0 to marginal calls. This matrix was clustered without further mathematical transformation. Only genes were clustered, i.e. the order of the samples is the order used on the Amazonia! website, and is based on grouping according to the embryonic germ layer origin of the sample.
hESC lines and karyotype
The list of hESC lines used and their respective karyotype are listed in Table 1.
Table 1.
hESC: laboratory of origin and karyotype
| hESC | Laboratory of origin | Karyotype |
|---|---|---|
| HUES1 | Douglas Melton, Harvard | p20 - 46,XX
p30 - 46,XX p40 - 46,XX, t(1;10) |
| HUES3 | Douglas Melton, Harvard | p23 - 46,XY
p49 - 47,XXY |
| HS293 | Outi Hovatta, Karolinska Institute | p49 - 46,XY |
| HS235 | Outi Hovatta, Karolinska Institute | p75 - 46,XX |
hESC culture
After approval from the French Ministry of Research, the French Ministry of Health and the Agence de la Biomédecine, HUES1 and HUES3 hESCs were imported from Douglas Melton’s laboratory (Harvard University, MA, USA) and cultured as described [24]. Cells were passaged every 3–4 days enzymatically with 0.25% trypsin/EDTA (Invitrogen, Cergy Pontoise, France) and cultured in knockout Dulbecco’s modified essential medium (Invitrogen) without plasmanate, with 10% KO-SR (Invitrogen), 2 mM L-glutamine, 1x nonessential amino acids, 0.05 mM β-mercaptoethanol, 10 ng/ml FGF2 (Abcys, Paris, France). Medium was replaced daily. HUES1 and HUES3 were cultured on murine embryonic fibroblasts (MEF) obtained from E13 ICR mice embryos (Harlan, Gannat, France) or on human foreskin fibroblasts (hFF) in porcine skin gelatin coated (Sigma-Aldrich, St. Quentin. Fallavier, France) 6 wells dishes. The hFF cell lines SM1 and SM3 were derived from respectively 75 and 3 years old patients undergoing foreskin reduction. Informed consent was obtained from the patient or the patient’s parents. MEF were cultured in DMEM with 10% fetal calf serum (FCS) and hFF in DMEM with 20% FCS. MEF and hFF were mitotically inactivated by mitomycin-C (2h at 10 μg/ml). These hESC expressed POU5F1, TRA-1-60, TRA-1-81, displayed phosphatase alkaline activity, and were able to differentiate into embryoid bodies that expressed differentiation markers of astrocytic lineage (GFAP) or endodermal lineage (alpha-feto protein) (see Supplemental Figure S1 and data not shown).
The HS293 and HS235 hESC lines were cultured in the Department of Obstetrics and Gynecology, (CLINTEC) at the Karolinska University Hospital as described [25]. Briefly, hESC were cultured on hFF (CRL-2429; American Type Culture Collection, Manassas, VA) mitotically inactivated by irradiation (40 Gy), in KO-DMEM 20% knockout SR, 2 mM Glutamax, 0.5% penicillin–streptomycin, 1% nonessential amino acids, 0.5 mM β-mercaptoethanol (all from Gibco Invitrogen Corporation, Paisley, Scotland), 1% insulintransferrin-selenium (Sigma-Aldrich) and 8 ng/mL of bFGF (R&D Systems, Oxford, UK). For karyotype analysis, hESC cells were treated with 75ng/ml final Karyomax Colcemid (Invitrogen) for 1h, trypsinized, incubated in 0.0375M KCl for 20 min, and fixed in fresh 3:1 methanol/acetic acid solution. 1 to 2 spreads were counted for chromosome number and 12 to 16 banding patterns were analyzed at 300–500 bands resolution.
Flow cytometry analysis
Human ES cells and fibroblasts were dissociated with trypsin (0.25%)-EDTA (1mM) (GIBCO) for 3 min. Cells were then washed with PBS and incubated for 30 min at 4°C in PBS with the corresponding monoclonal antibody (MAb): anti-CD24 MAb conjugated to phycoerythrin (PE) (dilution 1:50) (clone ALB9, Immunotech, Marseille, France) and/or anti-CD44 MAb conjugated to fluorescein isothiocyanate (FITC) (dilution 1:50) (clone J-173, Immunotech). After PBS washes, cells were suspended in Facsflow (Becton Dickinson, San Jose, CA) and fluorescence was analyzed with a FACSCalibur flow cytometer (Becton Dickinson) or sorted with a FACSAria cell sorter (Becton Dickinson). Appropriate isotype controls were included in all analyses.
Immunofluorescence
hESCs cultured on coverslips were fixed for 20 min in 4% paraformaldehyde and washed three times in PBS. Cells were permeabilized with 0.1% Triton X-100. After blocking at room temperature for 60 min in PBS with 5% donkey serum (S30, Chemicon international, Temecula, CA), cells were incubated for 1 hour at room temperature with primary antibody diluted in PBS with 5% donkey serum: POU5F1/OCT3/4 (sc 9081, Santa Cruz Biotechnology, Santa Cruz, CA; 1:300) and SEMA6A (AF1146, R&D, Abington, United Kingdom; 1:50). Cells were washed three times in PBS and incubated for 1 hour at room temperature with Alexa Fluor® 488 Donkey anti-Rabbit (A-11034; Molecular Probes; 1:1000) and Cy3 Donkey anti-Goat (Jackson ImmunoResearch; USA; 1:400) secondary antibodies, for POU5F1 and SEMA6A respectively. Unbound antibodies were removed by three washes in PBS. Hoechst staining was added to the first wash (Sigma, 5 μg/ml).
Results
Compiling hESC expression profiles
As of October 1st, 2006, we identified 38 original studies, 1protocol description and 4 reviews analyzing the transcriptome of hESCs (Table 2). The original studies used various hESCs, control cells and gene expression analysis techniques (summarized in Supplemental Table S1). 28 different hESC lines were used and transcriptome techniques included microarrays (7 different types of chips), ESTs scanning, SAGE, MPSS and Illumina beads. One study compared chromosome immuno-precipitation (ChIP) chip data with transcriptome data in hESCs [10]. Nevertheless, some common features emerged such as the frequent investigation of the H1, H9, and BG01 hESCs (in 12, 12 and 11 studies respectively), and the use of the GeneChip (Affymetrix) microarray system (in 16 studies) (see supplemental table S1).
Table 2.
43 original studies or reviews analyzing hESC transcriptome
| Reference | Technique/Review |
|---|---|
| (Abeyta et al., 2004) [43] | Microarray |
| (Aghajanova et al., 2006) [44] | Microarray |
| (Armstrong et al., 2006) [45] | Microarray |
| (Beqqali et al. 2006)[46] | Microarray |
| (Bhattacharya et al., 2004) [47] | Microarray |
| (Bhattacharya et al., 2005) [48] | Microarray |
| (Boyer et al., 2005) [10] | ChIP chip |
| (Brandenberger et al., 2004b) [30] | ESTs |
| (Brandenberger et al., 2004a) [49] | MPSS |
| (Brimble et al., 2004) [50] | MPSS |
| (Brimble et al. 2006) [51] | Microarray |
| (Cai et al., 2005) [52] | Illumina BeadChips |
| (Calhoun et al., 2004) [53] | Microarray |
| (Darr et al., 2006) [54] | Microarray |
| (Dvash et al., 2004) [55] | Microarray |
| (Enver et al., 2005) [56] | Microarray |
| (Ginis et al., 2004) [57] | Microarray |
| (Golan-Mashiach et al., 2004) [14] | Microarray |
| (Hirst et al.,2006) [19] | Microarray |
| (Josephson et al. 2006) [58] | Illumina BeadChips |
| (Kim et al., 2006) [59] | Microarray |
| (Li et al. 2006) [60] | Illumina BeadChips |
| (Liu et al. 2006) [61] | Illumina BeadChips |
| (Miura et al., 2004) [62] | MPSS and EST |
| (Perez-Iratxeta et al., 2005) [20] | Microarray |
| (Player et al., 2006) [63] | Microarray |
| (Pyle et al., 2004) [64] | Review |
| (Rao M., 2004) [65] | Review |
| (Rao R. et al., 2004) [66] | Microarray |
| (Rao R. and Stice, 2004) [67] | Review |
| (Richards et al., 2004) [68] | SAGE |
| (Richards et al., 2006) [69] | SAGE |
| (Robson, 2004) [70] | Review |
| (Sato et al., 2003) [15] | Microarray |
| (Sato and Brivanlou, 2006) [71] | Microarray (protocol description) |
| (Skottman et al., 2005) [16] | Microarray |
| (Skottman et al., 2006) [72] | Microarray |
| (Sperger et al., 2003) [29] | Microarray |
| (Suarez-Fariñas et al., 2005) [33] | Meta-analysis |
| (Sun et al., 2006) [73] | Microarray |
| (Wei et al., 2005) [31] | MPSS |
| (Xu et al. 2002) [74] | Microarray |
| (Zeng et al., 2004) [75] | Microarray |
Meta-analysis of genes differentially expressed between hESCs and non-pluripotent cells
One main objective of large scale gene expression analyses of hESCs is to identify the set of genes that are overexpressed in this unique cell type (“hESC” genes) or underexpressed (“differentiation” genes). We reasoned that bona fide “hESC” or “differentiation” genes would be repeatedly uncovered by independent groups, regardless of the hESC lines, the control cells, the assay format or the statistical method that had been used. We collected, from these 38 original studies, 20 lists of transcripts overexpressed in hESCs (“hESC genes”) and 11 lists of genes underexpressed in hESCs (“differentiation genes”).
The 20 lists of hESC genes comprised 5567 different genes. As illustrated in Figure 1A, we observed a marked heterogeneity between these lists, with only 1076 genes found overexpressed in hESCs by three or more independent studies, 48 genes by 10 or more studies and only 1 gene by all 20 lists (see Supplemental Table S2). The 48 genes found overexpressed in at least 10 studies are listed in Table 3. Of note, the pivotal ES transcription factor POU5F1/OCT3/4 is the one gene found by all 20 lists, whereas genes found in at least 10 studies include the transcription factors NANOG and SOX2, and the growth factors TDGF1/CRIPTO and Galanin (GAL) that are known to be highly expressed by hESCs. Thus, according to hESC transcriptome analyses published to date, this list of 1076 genes found overexpressed in hESC cells by three or more studies can be viewed as a “consensus hESC gene list”.
Figure 1. Meta-analysis of published “hESC” and “differentiation” gene lists.

1959 different genes were found overexpressed in hESCs compared to differentiated tissues by at least two out of 20 independent studies (purple pyramid) (A). Of these, 1076 genes were found by at least three studies, 48 genes by at least 10 studies and only 1 gene (POU5F1) by all studies. Eleven studies provided lists of genes downregulated in hESCs compared to more differentiated samples (yellow pyramid) (B). 1560 genes were found downregulated in hESC by at least two studies, 783 by at least 3 different studies, three by at least 9 different studies and none by all studies.
Heat map of gene expression detection for the hESC gene list across 24 hESC and 193 fetal and adult tissues samples analyzed with the U133A microarray (828 probesets) (C). Red stands for a “Present” detection call (i.e. gene expression confidently detected according to the GCOS 1.2 software), grey for “Marginal” and yellow for “Absent”. Cluster a: genes exclusively expressed in hESCs; b: gene expressed in hESCs and central nervous system samples; c: cell cycle genes; d: genes expressed in hESCs and in most tissues analyzed. The clustering was carried out using the Cluster and Treeview software. Only genes were clustered. hFF: human foreskin fibroblasts; OV: ovary; CNS: central nervous system; PNS: peripheral nervous system; SK: skin and keratinocytes; LU: lung; DT: digestive tract; TH: thyroid; AD: adipocytes; K&P: kidney and prostate; H&M: heart and muscle; HEMATO: various hematopoietic tissues; UT: uterus; PL: placenta.
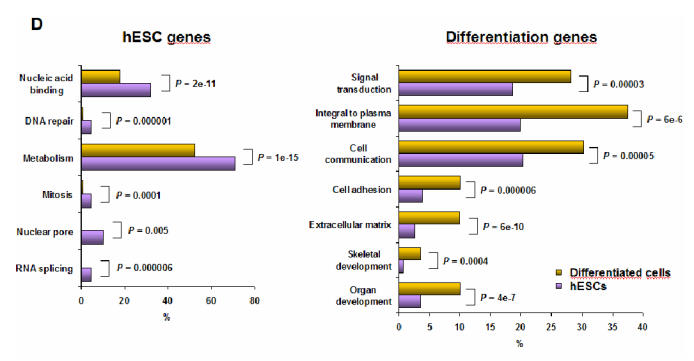
Comparison of GO annotations between hESC genes and differentiation genes (D). We compared the frequency of GO annotations of genes overexpressed in hESCs to those underexpressed in hESCs. The statistical analysis was carried out using the Babelomics webtool (http://babelomics.bioinfo.cipf.es/). Histograms show the percentage of genes with the specified GO annotation in the group of genes overexpressed in hESCs (purple) or in differentiated tissues (yellow). P : adjusted FDR P-value.
Table 3.
48 genes overexpressed in hESCs compared to differentiated cell types in at least 10 studies
| Abbreviation | Name | Chr.£ | Unigene | Nb of studies | Ratio Und/Dif Signal§ |
|---|---|---|---|---|---|
| POU5F1 | POU domain, class 5, transcription factor 1 | 6p21.31 | Hs.249184 | 20 | 56.4 |
| TDGF1 | Teratocarcinoma-derived growth factor 1 | 3p21.31 | Hs.385870 | 17 | 55.0 |
| DPPA4 | Developmental pluripotency associated 4 | 3q13.13 | Hs.317659 | 16 | 30.1 |
| LIN28 | Lin-28 homolog (C. elegans) | 1p36.11 | Hs.86154 | 16 | 24.8 |
| NANOG | Nanog homeobox | 12p13.31 | Hs.329296 | 15 | 88.9 |
| DNMT3B | DNA (cytosine-5-)-methyltransferase 3 beta | 20q11.2 | Hs.251673 | 15 | 27.8 |
| TERF1 | Telomeric repeat binding factor (NIMA-interacting) 1 | 8q13 | Hs.442707 | 15 | 18.9 |
| SEMA6A | Semaphorin 6A | 5q23.1 | Hs.156967 | 15 | 12.3 |
| M6PR | Mannose-6-phosphate receptor (cation dependent) | 12p13 | Hs.134084 | 15 | 10.6 |
| SNRPN | Small nuclear ribonucleoprotein polypeptide N | 15q11.2 | Hs.525700 | 15 | 7.3 |
| FLJ10884 | Hypothetical protein FLJ10884 | 1p31.3 | Hs.562195 | 14 | 260.7 |
| LEFTY1 | Left-right determination factor 1 | 1q42.1 | Hs.278239 | 14 | 34.1 |
| GAL | Galanin | 11q13.2 | Hs.278959 | 14 | 21.4 |
| SEPHS1 | Selenophosphate synthetase 1 | 10p14 | Hs.124027 | 14 | 6.3 |
| GABRB3 | Gamma-aminobutyric acid (GABA) A receptor, beta 3 | 15q11.2-q1 | Hs.302352 | 13 | 15.3 |
| SOX2 | SRY (sex determining region Y)-box 2 | 3q26.3-q27 | Hs.518438 | 13 | 15.3 |
| LECT1 | Leukocyte cell derived chemotaxin 1 | 13q14-q21 | Hs.421391 | 12 | 37.2 |
| LOC90806 | Similar to RIKEN cDNA 2610307I21 | 1q32.3 | Hs.157078 | 12 | 14.6 |
| BUB1 | BUB1 budding uninhibited by benzimidazoles 1 homolog | 2q14 | Hs.469649 | 12 | 11.2 |
| PSIP1 | PC4 and SFRS1 interacting protein 1 | 9p22.3 | Hs.493516 | 12 | 5.4 |
| INDO | Indoleamine-pyrrole 2,3 dioxygenase | 8p12-p11 | Hs.840 | 11 | 34.4 |
| HELLS | Helicase, lymphoid-specific | 10q24.2 | Hs.546260 | 11 | 19.3 |
| GPC4 | Glypican 4 | Xq26.1 | Hs.58367 | 11 | 15.4 |
| ITGB1BP3 | Integrin beta 1 binding protein 3 | 19p13.3 | Hs.135458 | 11 | 15.3 |
| CYP26A1 | Cytochrome P450, family 26, subfamily A, polypeptide 1 | 10q23-q24 | Hs.150595 | 11 | 14.2 |
| MCM5 | MCM5 minichromosome maintenance deficient 5 | 22q13.1 | Hs.517582 | 11 | 11.9 |
| MTHFD1 | Methylenetetrahydrofolate dehydrogenase 1 | 14q24 | Hs.435974 | 11 | 8.7 |
| PPAT | Phosphoribosyl pyrophosphate amidotransferase | 4q12 | Hs.331420 | 11 | 8.3 |
| SLC16A1 | AKR7 family pseudogene | 1p12 | Hs.75231 | 11 | 7.8 |
| NASP | Nuclear autoantigenic sperm protein (histone-binding) | 1p34.1 | Hs.319334 | 11 | 7.4 |
| DLG7 | Discs, large homolog 7 (Drosophila) | 14q22.3 | Hs.77695 | 11 | 6.7 |
| UGP2 | UDP-glucose pyrophosphorylase 2 | 2p14-p13 | Hs.516217 | 11 | 6.1 |
| MSH2 | MutS homolog 2, colon cancer, nonpolyposis type 1 | 2p22-p21 | Hs.156519 | 11 | 3.4 |
| ZIC3 | Zic family member 3 heterotaxy 1 | Xq26.2 | Hs.111227 | 10 | 30.7 |
| PIM2 | Pim-2 oncogene | Xp11.23 | Hs.496096 | 10 | 18.5 |
| HRASLS3 | HRAS-like suppressor 3 | 11q12.3-q1 | Hs.502775 | 10 | 16.8 |
| ALPL | Alkaline phosphatase, liver/bone/kidney | 1p36.1-p34 | Hs.75431 | 10 | 15.0 |
| AASS | Aminoadipate-semialdehyde synthase | 7q31.3 | Hs.433075 | 10 | 13.8 |
| MGST1 | Microsomal glutathione S-transferase 1 | 12p12.3-p1 | Hs.389700 | 10 | 10.9 |
| GDF3 | Growth differentiation factor 3 | 2p13.1 | Hs.86232 | 10 | 9.5 |
| HSPA4 | Heat shock 70kDa protein 4 | 5q31.1-q31 | Hs.90093 | 10 | 9.2 |
| PHF17 | PHD finger protein 17 | 4q26-q27 | Hs.12420 | 10 | 9.2 |
| UNG | Uracil-DNA glycosylase | 12q23-q24. | Hs.191334 | 10 | 6.7 |
| USP9X | Ubiquitin specific peptidase 9, X-linked | Xp11.4 | Hs.77578 | 10 | 5.7 |
| CEBPZ | CCAAT/enhancer binding protein zeta | 2p22.2 | Hs.135406 | 10 | 5.4 |
| CRMP1 | Collapsin response mediator protein 1 | 4p16.1-p15 | Hs.135270 | 10 | 5.3 |
| BMPR1A | Bone morphogenetic protein receptor, type IA | 10q22.3 | Hs.524477 | 10 | 5.1 |
| ORC1L | Origin recognition complex, subunit 1-like (yeast) | 1p32 | Hs.17908 | 10 | 3.9 |
Chromosomal location.
Ratio of mean expression in hESC samples to mean expression in non-hESC samples.
In order to get further insights into this hESC list, we built an expression compendium by combining the data from 5 publications using the U133A GeneChip microarray to analyze hESC transcriptome and 3 publications providing the transcriptome of various normal fetal and adult tissues (see material and method and Supplemental Table S4). This compendium included the gene profiling of 24 hESC samples and more than 190 various fetal and adult tissues samples. A heat map was generated for the “consensus hESC gene list” in this expression compendium based on the detection call provided by the GCOS 1.2 software. The detection call is a way to evaluate whether a gene is expressed or not in a given sample. Hierarchical clustering (Figure 1C) delineated four major clusters of genes: cluster (a) was a group of 40 genes specifically detected in hESCs (“hESC specific” genes, Table 4), including expected genes such as POU5F1/OCT3/4, NANOG, TDGF1, LIN28, CLDN6, GDF3, DNMT3A, but also genes such as CYP26A1, HELLS or GPR19, cluster (b) featured genes that were detected in both hESCs and CNS samples such as the GABA receptors GABRB3 and GABRA5, and the growth factor FGF13, cluster (c) genes detected in samples characterized by a high mitotic index such as SKP2, MYC, the cyclin CCNA2 and the MCM genes MCM2, MCM5, MCM6 and MCM7, cluster (d) genes overexpressed in hESCs but also expressed in a majority of the tissues included in this dataset such as PGK1, HSPA9B, and the ribosomal genes RPLP0, RPL6, RPL7 and RPL24. The complete lists of genes composing these clusters are available as Supplemental Table S5. The expression histograms of the 40 hESC specific genes are shown in Supplemental Figure S2. These results show that, though all 1076 genes of the “consensus hESC gene list” have been found overexpressed in hESCs compared to non-hESC samples by at least three different studies, 40 genes are indeed hESC specific (cluster a) but most are nonetheless also expressed in adult tissues to various extent.
Table 4.
40 genes specifically expressed in hESCs (cluster “a” in Figure 1C)
| Abbreviation | Name | Chr.£ | Unigene | Nb of studies | Ratio Und/Dif Signal § | Presence Call in non-hESC tissues # |
|---|---|---|---|---|---|---|
| NANOG | Nanog homeobox | 12p13.31 | Hs.329296 | 15 | 88.9 | 0 |
| POU5F1 | POU domain, class 5, transcription factor 1 | 6p21.31 | Hs.249184 | 20 | 56.4 | 0 |
| LIN28 | lin-28 homolog (C. elegans) | 1p36.11 | Hs.86154 | 16 | 24.8 | 0 |
| CDC25A | cell division cycle 25A | 3p21 | Hs.1634 | 5 | 19.4 | 0 |
| PRDM14 | PR domain containing 14 | 8p21-p12 | Hs.287532 | 6 | 13.5 | 0 |
| SLD5 | SLD5 homolog | 8p11.21 | Hs.521557 | 5 | 13.2 | 0 |
| C14orf115 | chromosome 14 open reading frame 115 | 14q24.3 | Hs.196530 | 9 | 12.9 | 0 |
| GDF3 | growth differentiation factor 3 | 12p13.1 | Hs.86232 | 10 | 9.5 | 0 |
| GPR19 | G protein-coupled receptor 19 | 12p12.3 | Hs.92458 | 6 | 6.3 | 0 |
| GPR23 | G protein-coupled receptor 23 | Xq13-q21.1 | Hs.522701 | 3 | 2.7 | 0 |
| FLJ10884 | hypothetical protein FLJ10884 | 1p31.3 | Hs.562195 | 14 | 260.7 | 1 |
| ZIC3 | Zic family member 3 heterotaxy 1 | Xq26.2 | Hs.111227 | 10 | 30.7 | 1 |
| CLDN6 | claudin 6 | 16p13.3 | Hs.533779 | 9 | 14.4 | 1 |
| CYP26A1 | cytochrome P450, family 26, A1 | 10q23-q24 | Hs.150595 | 11 | 14.2 | 1 |
| CXorf15 | chromosome X open reading frame 15 | Xp22.2 | Hs.555961 | 3 | 12.4 | 1 |
| HESX1 | homeo box (expressed in ES cells) 1 | 3p21.2-p21 | Hs.171980 | 5 | 10.3 | 1 |
| KIAA0523 | KIAA0523 protein | 17p13.2 | Hs.370166 | 4 | 6.5 | 1 |
| MGC3101 | hypothetical protein MGC3101 | 16q24.3 | Hs.301394 | 5 | 5.0 | 1 |
| CHEK2 | CHK2 checkpoint homolog (S. pombe) | 22q11 | Hs.558896 | 7 | 3.3 | 1 |
| RNU3IP2 | RNA, U3 small nucleolar interacting protein 2 | 3p21.2 | Hs.153768 | 4 | 3.0 | 1 |
| GJA7 | gap junction protein, alpha 7 | 17q21.31 | Hs.532593 | 4 | 2.3 | 1 |
| ETV4 | ets variant gene 4 | 17q21 | Hs.434059 | 9 | 9.8 | 2 |
| DNMT3A | DNA (cytosine-5-)-methyltransferase 3 alpha | 2p23 | Hs.515840 | 5 | 4.2 | 2 |
| DCC1 | defective in sister chromatid cohesion homolog 1 | 8q24.12 | Hs.315167 | 5 | 3.5 | 2 |
| SLC5A6 | solute carrier family 5, member 6 | 2p23 | Hs.435735 | 3 | 2.9 | 2 |
| TDGF1 | teratocarcinoma-derived growth factor 1 | 3p21.31 | Hs.385870 | 17 | 55.0 | 3 |
| PWP2H | PWP2 periodic tryptophan protein homolog | 21q22.3 | Hs.449076 | 4 | 27.0 | 3 |
| ABHD9 | abhydrolase domain containing 9 | 19p13.12 | Hs.156457 | 3 | 9.7 | 3 |
| EPHA1 | EPH receptor A1 | 7q34 | Hs.89839 | 7 | 8.0 | 3 |
| MYBL2 | v-myb homolog-like 2 | 20q13.1 | Hs.179718 | 3 | 4.0 | 3 |
| PRO1853 | hypothetical protein PRO1853 | 2p22.2 | Hs.433466 | 5 | 5.9 | 4 |
| ORC1L | origin recognition complex, subunit 1-like (yeast) | 1p32 | Hs.17908 | 10 | 3.9 | 4 |
| MCM10 | minichromosome maintenance deficient 10 | 10p13 | Hs.198363 | 5 | 3.8 | 4 |
| FLJ20105 | FLJ20105 protein | Xq13.1 | Hs.47558 | 5 | 3.3 | 4 |
| ORC2L | origin recognition complex, subunit 2-like (yeast) | 2q33 | Hs.444870 | 3 | 2.6 | 4 |
| FLJ12484 | interferon stimulated exonuclease like 1 | 15q26.1 | Hs.37747 | 6 | 4.0 | 5 |
| HELLS | helicase, lymphoid-specific | 10q24.2 | Hs.546260 | 11 | 19.3 | 6 |
| BRRN1 | barren homolog (Drosophila) | 2q11.2 | Hs.308045 | 5 | 4.6 | 6 |
| RBM14 | RNA binding motif protein 14 | 11q13.2 | Hs.11170 | 3 | 4.1 | 7 |
| DTYMK | deoxythymidylate kinase (thymidylate kinase) | 2q37.3 | Hs.471873 | 3 | 3.1 | 7 |
Chromosomal location.
Ratio of mean expression in hESC samples to mean expression in non-hESC samples.
number of non-hESC samples with a detection call “Present” (out of 193 non-hESC samples).
The 11 lists of “differentiation” genes summed up to 4798 genes and we noted a similar heterogeneity as in the “hESC” lists (Supplemental Table S3). Out of these 4798 genes, only 783 were found underexpressed in hESCs by at least 3 different studies, composing a “consensus differentiation gene list”, three genes, lumican, collagen 1A1 and 3A1, by nine studies and none by all 11 studies (Figure 1B). Table 5 shows 30 selected genes found by at least 6 studies, which include the bone morphogenetic proteins (BMP) 1 and 4, the keratins 7 and 18, insulin-like growth factor 2 (IGF2), Heart and neural crest derivatives expressed 1 (HAND1) and the transducing chain IL6ST/GP130. The complete “consensus differentiation gene list” can be found in Supplemental Table S3.
Table 5.
30 selected genes underexpressed in hESCs compared to differentiated cell types and found by at least 6 studies
| Abbreviation | Name | Unigene | Chr.£ | Nb of studies | Ratio Dif/Und Signal § |
|---|---|---|---|---|---|
| LUM | Lumican | Hs.406475 | 12q21.3 | 9 | 18.1 |
| COL3A1 | Collagen, type III, alpha 1 | Hs.443625 | 2q31 | 9 | 14.0 |
| COL1A1 | Collagen, type I, alpha 1 | Hs.172928 | 17q21.3 | 9 | 5.8 |
| IGFBP3 | Insulin-like growth factor binding protein 3 | Hs.450230 | 7p13-p12 | 8 | 14.1 |
| ACTA2 | Actin, alpha 2, smooth muscle, aorta | Hs.500483 | 10q23.3 | 8 | 14.0 |
| P4HA2 | Procollagen-proline, 2-oxoglutarate 4-dioxygenase A2 | Hs.519568 | 5q31 | 8 | 5.5 |
| SPARC | Secreted protein, acidic, cysteine-rich (osteonectin) | Hs.111779 | 5q31.3 | 8 | 4.7 |
| COL1A2 | Collagen, type I, alpha 2 | Hs.489142 | 7q22.1 | 8 | 4.6 |
| KRT18 | Keratin 18 | Hs.406013 | 12q13 | 8 | 3.8 |
| DCN | Decorin | Hs.156316 | 12q13.2 | 7 | 39.9 |
| BMP4 | Bone morphogenetic protein 4 | Hs.68879 | 14q22 | 7 | 15.0 |
| COL5A1 | Collagen, type V, alpha 1 | Hs.210283 | 9q34 | 7 | 12.6 |
| COL2A1 | Collagen, type II, alpha 1 | Hs.408182 | 12q13 | 7 | 10.5 |
| RHOBTB3 | Rho-related BTB domain containing 3 | Hs.445030 | 5q15 | 7 | 6.4 |
| NDRG1 | N-myc downstream regulated gene 1 | Hs.372914 | 8q24.3 | 7 | 6.1 |
| COL6A3 | Collagen, type VI, alpha 3 | Hs.233240 | 2q37 | 7 | 6.0 |
| CST3 | Cystatin C | Hs.304682 | 20p11.21 | 7 | 5.3 |
| KRT8 | Keratin 8 | Hs.533782 | 12q13 | 7 | 5.2 |
| CD47 | CD47 antigen (Rh-related antigen) | Hs.446414 | 3q13.1 | 7 | 3.5 |
| HAND1 | Heart and neural crest derivatives expressed 1 | Hs.152531 | 5q33 | 6 | 61.9 |
| IGF2 | Insulin-like growth factor 2 (somatomedin A) | Hs.373908 | 11p15.5 | 6 | 23.6 |
| CXCL14 | Chemokine (C-X-C motif) ligand 14 | Hs.483444 | 5q31 | 6 | 10.5 |
| COL11A1 | Collagen, type XI, alpha 1 | Hs.523446 | 1p21 | 6 | 8.7 |
| IGFBP7 | Insulin-like growth factor binding protein 7 | Hs.479808 | 4q12 | 6 | 8.0 |
| IL6ST | Interleukin 6 signal transducer (gp130) | Hs.532082 | 5q11 | 6 | 7.6 |
| COL5A2 | Collagen, type V, alpha 2 | Hs.445827 | 2q14-q32 | 6 | 6.8 |
| KRT7 | Keratin 7 | Hs.411501 | 12q12 | 6 | 6.5 |
| KRT19 | Keratin 19 | Hs.514167 | 17q21.2 | 6 | 6.5 |
| BMP1 | Bone morphogenetic protein 1 | Hs.1274 | 8p21 | 6 | 6.1 |
| CDKN1A/P21 | Cyclin-dependent kinase inhibitor 1A (p21, Cip1) | Hs.370771 | 6p21.2 | 6 | 4.6 |
Chromosomal location.
Ratio of mean expression in non-hESC samples to mean expression in hESC samples.
We compared functional Gene Ontology (GO) annotations of the hESC genes to the differentiation genes. Several functional annotations were more represented in each category (Figure 1D). There were significantly more genes involved in “metabolism”, “mitosis”, “RNA splicing”, “nuclear pore”, “DNA repair” in the hESC gene list, reflecting the intense proliferation, DNA replication and DNA remodeling taking place in these cells. Conversely, GO annotations such as “organ development”, “skeletal development”, “extracellular matrix”, “cell adhesion”, “cell communication”, “integral to plasma membrane”, “signal transduction”, were significantly more frequent among genes upregulated in differentiated tissues, in agreement with the idea that hESC differentiation mimics early organogenesis, which is associated with the development of complex cell-cell communication and cell-extra cellular matrix (ECM) interactions.
CD24 and SEMA6A are hESC markers
We next inquired whether we could find, among the hESC genes, new markers that may be useful to identify, isolate and qualify hESCs in vitro. We focused on two cell surface hESC genes: CD24 and SEMA6A. These two genes have been found to be overexpressed in hESCs by respectively nine and fifteen studies, and are therefore good candidates as new hESC markers.
CD24 is a sialoglycoprotein known to be expressed on mature granulocytes and B cells subpopulations [26]. In addition to these hematopoietic cell types, microarray data from the expression compendium evidenced a high CD24 mRNA expression in keratinocytes, pancreas and thyroid, whereas most neural tissues, muscle, liver and testis did not express CD24 (Figure 2A). Upon differentiation of hESCs into embryoid bodies (EBs), CD24 expression is markedly downregulated (Figure 2B). Importantly, human foreskin fibroblasts (hFF) samples did not express CD24 (Figure 2C, 2F). Hence, we investigated whether CD24 could discriminate hESCs from fibroblasts in culture. We analyzed by flow cytometry the hESC lines HUES1, HUES3, HS293 and HS235 [24, 25] cultured on hFF and evidenced two distinct cell populations, one CD24+ and one CD24− (Figure 2E). To demonstrate that the CD24+ population corresponded to hESCs and the CD24− population to hFF, we took advantage of CD44, a strong fibroblast marker not expressed by hESCs (Figures 2D, 2G). Double staining of hESCs cultured on hFF showed that these two markers were expressed in a mutually exclusive manner and delimited a hESC CD24+CD44− population that did not overlap with the fibroblast CD24−CD44+ population (Figure 2H). Using CD24/CD44 labeling, we were able to separate hESC from fibroblasts and obtain pure hESC populations that recovered the cell sorting procedure and grew in vitro while retaining POU5F1 and cardinal cell surface markers of pluripotency expression (Supplemental Figure S3 and data not shown).
Figure 2. CD24 and SEMA6A are two new hESC markers.


Expression histograms of CD24 across the U133A compendium (A). These histograms were obtained from our Amazonia! website from the U133A oligonucleotide microarray (Affymetrix) datasets. The Y-axis features the signal value obtained with GCOS 1.2 software. Microarray expression histograms of CD24 gene before and after non-lineage differentiation into EBs (data from the study of [56]) (B). Mean U133A microarray signal value of CD24 (C) and CD44 (D) in 24 hESC samples versus two human fibroblasts samples. Flow cytometry analysis of CD24 on HUES1 hESCs co-cultured with hFF (E), of hFF alone (F) and of CD44 on hFF alone (G), and double CD24 and CD44 labeling of HUES1 (passage 37), HUES3 (p43), HS235 (p30) and HS293 (p68) cells co-cultured with hFF (H). Expression histogram of SEMA6A (I). Microarray expression histograms of SEMA6A gene before and after differentiation into EBs (data from the study of [56]) (J). Mean U133A microarray signal value of SEMA6A in 24 hESC samples versus two human fibroblasts samples (K). Indirect immunofluorescence localization of SEMA6A, POU5F1/OCT3/4, or both and corresponding Hoechst nuclear staining in the HUES1 (L) and HUES3 hESCs (M) cultured on murine embryonic and human fibroblasts, respectively. Scale bar: 25μm. White arrow: MEF (L) or hFF (M).
SEMA6A is a class 6 semaphorin [27], i.e. transmembrane with cytoplasmic domain, known to be expressed in developing neural tissue. We recently reported the expression of this semaphorin in cumulus oophorus cells [28]. Comparison of RNA expression in hESCs and normal adult tissues showed that in addition to hESCs, SEMA6A was also expressed at high level in adult samples from the central and peripheral nervous system and placenta (Figure 2I). As for CD24, SEMA6A mRNA is downregulated upon EB differentiation and is not detected in human fibroblasts (Figure 2J and K). Immunofluorescence analysis showed a membrane localization of SEMA6A on hESCs, in contrast to POU5F1/OCT3/4 which had a strict nuclear localization (Figure 2L-M). In summary, we showed that CD24 and SEMA6A have indeed a preferential expression in hESCs, that this expression is confirmed at the protein level, and is declining upon EB differentiation. Thus these markers can be used to discriminate hESCs from feeder fibroblasts
hESC transcriptome data visualization through an open access web interface
We developed a website, Amazonia! (http://amazonia.montp.inserm.fr), to allow the scientific community access to these public data. This website is dedicated to the visualization of large, publicly available, human transcriptome data (Le Carrour et al., manuscript submitted). A main topic of this website is human embryonic stem cells. Data are visualized as expression histograms with a color code facilitating the recognition of cell type. Genes are accessed either by key words or through lists of genes. Most interestingly, when data were obtained using the same platform format, sample labeling and data normalization, it was possible to combine different experiments in one single virtual experiment. The U133A expression compendium comprises for example more than 200 different samples from 8 publications including hESCs and normal adult tissues. Thus, Amazonia! provides the expression profile of about 15 000 different genes in about 100 different tissues types or purified cell populations including 11 hESCs (H1, H9, HS181, HS235, HS237, FES21, FES22, FES29, FES30, ,I6, HES-2). Figure 3A illustrates this feature of our website with the expression histograms of five hESC specific genes (POU5F1, NANOG, GPR19, Helicase lymphoid-specific (HELLS) and the cytochrome P450 CYP26A1), two genes expressed in nonlineage-differentiated cells (HAND1 and IGF2), two factors highly expressed by human fibroblasts and smooth muscle that may contribute to the supporting properties of fibroblast to hESC culture (Gremlin (GREM1) and Matrix metalloproteinase 1 (MMP1)), one hematopoietic marker (CD45), one central nervous system marker (Glial fibrillary acidic protein (GFAP)) and one ubiquitously expressed gene (Ribosomal protein L3 (RPL3)).
Figure 3. Expression of selected genes using Amazonia!


Expression histograms of five hESC specific genes (POU5F1, NANOG, GPR19, HELLS and CYP26A1), two genes expressed in nonlineage-differentiated hESC cells (HAND1 and IGF2), two factors highly expressed by human fibroblasts and smooth muscle that may contribute to the supporting properties of fibroblasts to hESC culture (Gremlin and MMP1), one hematopoietic marker (CD45), one central nervous system specific gene (GFAP) and one ubiquitously expressed gene (Ribosomal protein L3 (RPL3))(A).
FZD7 expression in three distinct datasets using the Amazonia! web tool: in the “Embryonic and adult normal tissues” series (Affymetrix U133A microarray, from 8 different publications) (B), in the “hESC” series from Enver et al. (Affymetrix U133A) [56] (C) and in the “hESC, germinal and embryonic malignancies” series (Standford microarray) [29]. EC: embryonic carcinoma; YSC: Yolk sac carcinoma. (D)
Comparison of the expression of CD24 mRNA across normal (NL) and malignant lung (ML) samples (E), and across normal hematopoietic (NH) and B acute lymphoblastic leukemia (ALL) and T ALL samples (F).
Another important feature of Amazonia! is the possibility to compare on the same web page the expression of a gene of interest in various datasets. Figure 3B shows that the expression of Frizzled 7 (FZD7) was markedly upregulated in hESCs, was downregulated during non-lineage differentiation into EBs, and was also highly expressed in embryonal carcinoma and Yolk sac carcinoma samples. The combination of these three histograms evidences a preferential FZD7 mRNA expression in normal and malignant embryonic cells, suggesting that FZD7 may play a major role in these pluripotent cell types.
In order to facilitate access for the scientific community to the lists of genes from published transcriptome analyses, we implemented a list manager in our Amazonia! website. Thus, one can access these lists straightforwardly and obtain an expression histogram for each gene in various public transcriptome collections. This feature is of particular interest to challenge a list of genes, for example a “hESC” gene list, with other hESC datasets or even with non-hESC datasets such as cancer datasets. Indeed, once a gene is selected, the user can navigate between various thematic pages, switching for instance from the “stem cells” page to the “leukemia and lymphoma”, “lung cancer” or other pages. Using this feature of the Amazonia! website, we could observe that the sialoglycoprotein CD24 was also highly expressed in acute lymphoblastic leukemia, lung cancer and glioma samples when compared to the corresponding normal samples (Figure 3C and data not shown).
Discussion
Human ESCs are remarkable by their ability both to self renew and to generate virtually any kind of cell type, hence carrying many hopes for cell therapy. It is anticipated that genome wide expression analyses, by providing an extensive molecular taxonomy, will help understanding this unique cell model of stem cell pluripotency. Transcriptome results can be viewed as a non-biased, genome wide, expression catalog. Many groups have published the transcriptome of hESCs, but, as a direct consequence of the massive data generated, access to this data on a routine basis was precluded for most researchers. Therefore, the construction of a database collecting publicly available hESC transcriptome and accessible through a user friendly interface is of utmost interest for the hECS researchers’ community. We found 38 original studies or reviews analyzing the transcriptome of hESCs. Expression data and gene lists extracted from these studies were included into Amazonia! and are now readily accessible. Interestingly, the frequent use of the U133A oligonucleotide microarrays allowed us to construct a virtual expression dataset of about 15 000 different genes in more than 200 tissue samples from various origins, including 24 hESC samples. Hence, the expression of each gene in hESCs is directly contrasted to that of normal fetal and adult tissues, as illustrated in Figure 3A.
Unearthing the hESC gene panoply may help to define what makes hESCs unique. To achieve this goal, most studies compared hESC transcriptome data to that of more differentiated cell types and obtained lists of genes over- or underexpressed in hESCs. Comparison of different transcriptome surveys of hESCs gave us the unique opportunity to identify genes that were identified by several authors as differentially expressed in hESCs. However, a striking heterogeneity was observed between the 20 lists of hESC genes, and only 48 genes were found to be highly enriched in hESCs in at least ten or publications, among 5567 genes found in at least one. Some of these differences may be explained by platform-to-platform or lab-to-lab variability, but this is likely not the main explanation as suggested by transcriptome platform comparisons [22]. Rather, the differences between the hESC cells lines used, the control samples, the specific caveats of each transcriptome analysis technique and the statistical methodologies likely contribute to these disparities (see Supplemental Table S2). For instance, the homeobox transcription factor NANOG, which is universally expressed in hESC, was not reported by Sperger et al. because no probe for NANOG was present on their microarray [29], nor was it reported by Brandenberger et al. because the differential regulation of NANOG in their in vitro differentiation model did not met their stringent statistical criteria [30]. ZFP42 (the human homolog of murine rex1) was never listed by MPSS studies because its MPSS signature has repeat sequences [31]. Another pitfall impacting on differentially expressed genes lists comparisons and contributing to the “small intersection” problem [32] is that in order to be at the intersection of 20 lists, a gene must have fulfilled 20 times the statistical filter, which it does with a probability equal to the product of the probabilities of each test. A way to circumvent this difficulty would be to obtain the raw data from these studies and to apply specific statistical tests [33]. However, the raw data were not available for many studies analyzed here, which prevented us from applying this approach in our study. Nevertheless, the 1076 hESC genes list provides the opportunity to the scientific community to examine the genes that have found over- or underexpressed in hESC by several authors. These lists provide further molecular insight into the biology of this unique stem cell model and are now starting points for many new research directions in the field of hESC. In the future, it will be interesting to extend this list by investigating additional hESC lines. The collection of additional transcriptome data is ongoing and we will update the database according to new publications on the hESC genome expression.
As one can easily notice by brownsing the “consensus hESC gene list” in the U133A “embryonic and adult samples” dataset, we found very few genes that are completely hESC specific. Indeed, most genes found overexpressed in hESCs are also expressed in other tissues (Figure 1C), and only 40 genes, grouped in cluster (a), comprised genes which expression was not detected in most other tissues (see supplemental Figure S2). Of note, genes can only be labeled as hESC “specific” in respect of the different tissues and cell populations that have been tested. As new cell types will be investigated, some of these genes may be found expressed in these new samples and thereby cease to be specific to hESCs. Specificity also depends on the sensitivity of the assay investigating expression of a given gene. For example, POU5F1/OCT3/4 has been reported to be expressed in germinal cells and even in bone marrow by RT-PCR assays [34, 35]. By contrast, microarray analysis show that this gene, clearly expressed at a high level in hESCs and teratocarcinoma samples, is neither detected by this technique in testis, ovary, bone marrow samples, nor in a pure oocyte population (Figure 3A and unpublished data). These observations suggest that the properties of hESCs, comprising self-renewal, unrestrained proliferation and pluripotency, are mediated by the expression of few specific genes, if any, together with genes which individually are not hESC specific, but whose combined expression is specific to hESCs. The “consensus hESC gene list” should encompass many of those genes contributing to the embryonic stem cell characteristics.
Transcriptome approaches have several limitations. Each technique has its own technical limits, but this meta-analysis partly circumvented these by the simultaneous analysis of different, complementary methods. Another drawback is that by looking at the transcriptome, we consider gene expression at the RNA level only, eluding all forms of post-transcriptional regulation. It will therefore be important in the future to look the differential expression of the genes described in the hESC and differentiation lists at these hESC and differentiation gene lists at the protein level. For that matter, this meta-analysis provides a list of pertinent genes for further protein validation. In line with this proposition, we chose to investigate more thoroughly two cell surface protein-encoding genes, which may serve as new hESC markers: CD24 and SEMA6A. Though these two genes have been found to be overexpressed in hESCs by 9 and 15 studies, respectively, we found that hESCs shared this RNA expression with several adult tissues as substantiated by microarray results (Figure 2). However, purification of hESCs requires only distinguishing them from differentiating stem cells and from the co-cultured feeder cells. CD24 expression is low in differentiating hESCs, and is absent in human fibroblasts. We were thus able to purify by flow cytometry pure hESC populations. This provides a new tool to isolate highly enriched populations of hESCs for subsequent experiments, including microarray analysis. We observed that CD24 was also highly expressed on various malignant cell types, as previously reported [36, 37]. Since CD24 is a ligand for P-selectin, it has been suggested that CD24 could be important in the dissemination of tumor cells by facilitating the interaction with endothelial cells [38]. The role of CD24 in hESCs is less clear because P-selectin is not expressed by human fibroblasts nor by hESCs themselves (data not shown), suggesting that CD24 may have other molecular functions. Semaphorins have been initially identified for their role in neuronal guidance as chemorepellents, but it is becoming clear that this large family of genes also plays important roles in organogenesis, vascularization, angiogenenesis and B lymphocyte signaling. We show here a clear protein expression of semaphorin 6A in hESCs, with an overexpression in hESCs compared to many other cells types, suggesting a functional role for this transmembrane molecule in cell-to-cell interaction or signaling in hESCs.
The clustering of the “consensus hESC gene list” based on the “detection call” identified a cluster of 40 genes with a high specificity in hESCs, with no expression in most samples from more than 100 different fetal and adult cell tissues (Table 4). In addition to genes clearly expected such as POU5F1/OCT3/4, NANOG or TDGF1, this cluster included genes whose hESC specificity had been overlooked. For example the expression of the G protein-coupled receptor 19 (GPR19) is restricted to hESCs and, if functional, may offer a possibility for in vitro intervention on proliferation or pluripotency of hESCs. Other hESC specific genes comprise the cytochrome P450 CYP26A1 which is responsible for retinoic acid degradation [39] or the helicase HELLS which is expressed at a moderate level in a few lymphoid samples but most importantly in hESCs and could be involved in DNA strand separation, including replication, repair, recombination, and transcription [40] (Figure 3A). Our meta-analysis also spotted additional interesting genes such as FZD7 which was identified as the frizzled receptor preferentially expressed on hESCs. Based on this expression, we hypothesize that FZD7 could be a major WNT receptor in hESCs. Thus, FZD7 could contribute to the pluripotency signal mediated by WNT previously reported in hESCs [41]. Regarding the “consensus differentiation gene list”, many genes are clearly related to differentiation such as collagen and keratin genes. We note that the transducing chain IL6ST/GP130, which is necessary to convey the signals from the IL-6 growth factor family including LIF, is expressed at a very low level in hESCs as compared to most differentiated adult tissues. This is in line with the largely accepted view that LIF signaling is dispensable for pluripotency in hESCs [2].
Conclusion
We analyzed 38 publications studying the hESC transcriptome. We propose a “consensus hESC gene list” and a “consensus differentiation gene list” that identifies the genes found respectively up- or downregulated in hESCs compared to differentiated samples by at least three publications. We provide the first tool to directly visualize the expression of most human genes in hESCs, and provide direct comparison with their expression in many normal and malignant tissues. This tool may be considered as the first hESC expression Atlas online. By providing an easy access to this large public data, we hope that Amazonia! will help boosting the translation of these invaluable expression information into biological applications.
Supplementary Material
Acknowledgments
We are grateful to the various labs that gave free access to their complete transcriptome data, in agreement with the MIAME recommendations [42]. We thank Ned Lamb and Cyril Berthenet for support with informatics (IGH de Montpellier), and Hassan Boukhaddaoui for cell imaging (Montpellier RIO Imaging). We are grateful to Isabelle Rodde-Astier, Antoine Héron and Valérie Duverger (MacoPharma) for decisive support for this project. We thank Geneviève Lefort, Marie Ponset and Franck Pellestor for assistance in hESC karyotyping.
References
- 1.Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature. 1981;292:154–156. doi: 10.1038/292154a0. [DOI] [PubMed] [Google Scholar]
- 2.Thomson JA, Itskovitz-Eldor J, Shapiro SS, et al. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282:1145–1147. doi: 10.1126/science.282.5391.1145. [DOI] [PubMed] [Google Scholar]
- 3.Chambers I, Colby D, Robertson M, et al. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell. 2003 Jun 30;113:643–655. doi: 10.1016/s0092-8674(03)00392-1. [DOI] [PubMed] [Google Scholar]
- 4.Nichols J, Zevnik B, Anastassiadis K, et al. Formation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4. Cell. 1998 Nov 30;95:379–391. doi: 10.1016/s0092-8674(00)81769-9. [DOI] [PubMed] [Google Scholar]
- 5.Hanna LA, Foreman RK, Tarasenko IA, Kessler DS, Labosky PA. Requirement for Foxd3 in maintaining pluripotent cells of the early mouse embryo. Genes Dev. 2002 Nov 15;16:2650–2661. doi: 10.1101/gad.1020502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rogers MB, Hosler BA, Gudas LJ. Specific expression of a retinoic acid-regulated, zinc-finger gene, Rex-1, in preimplantation embryos, trophoblast and spermatocytes. Development Dec. 1991;113:815–824. doi: 10.1242/dev.113.3.815. [DOI] [PubMed] [Google Scholar]
- 7.Graham V, Khudyakov J, Ellis P, Pevny L. SOX2 functions to maintain neural progenitor identity. Neuron. 2003 Aug 28;39:749–765. doi: 10.1016/s0896-6273(03)00497-5. [DOI] [PubMed] [Google Scholar]
- 8.Feldman B, Poueymirou W, Papaioannou VE, DeChiara TM, Goldfarb M. Requirement of FGF-4 for postimplantation mouse development. Science. 1995 Feb 13;267:246–249. doi: 10.1126/science.7809630. [DOI] [PubMed] [Google Scholar]
- 9.Lee TI, Jenner RG, Boyer LA, et al. Control of developmental regulators by polycomb in human embryonic stem cells. Cell. 2006 Apr 21;125:301–313. doi: 10.1016/j.cell.2006.02.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Boyer LA, Lee TI, Cole MF, et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell. 2005 Sep 23;122:947–956. doi: 10.1016/j.cell.2005.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rhodes DR, Yu J, Shanker K, et al. ONCOMINE: a cancer microarray database and integrated data-mining platform. Neoplasia Jan-Feb. 2004;6:1–6. doi: 10.1016/s1476-5586(04)80047-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Al-Shahrour F, Minguez P, Vaquerizas JM, Conde L, Dopazo J. BABELOMICS: a suite of web tools for functional annotation and analysis of groups of genes in high-throughput experiments. Nucleic Acids Res. 2005 Jul 1;33:W460–464. doi: 10.1093/nar/gki456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Su AI, Wiltshire T, Batalov S, et al. A gene atlas of the mouse and human protein-encoding transcriptomes. Proc Natl Acad Sci U S A. 2004 Apr 20;101:6062–6067. doi: 10.1073/pnas.0400782101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Golan-Mashiach M, Dazard JE, Gerecht-Nir S, et al. Design principle of gene expression used by human stem cells: implication for pluripotency. Faseb J. 2004 Oct 21; doi: 10.1096/fj.04-2417fje. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 15.Sato N, Sanjuan IM, Heke M, Uchida M, Naef F, Brivanlou AH. Molecular signature of human embryonic stem cells and its comparison with the mouse. Dev Biol. 2003 Aug 15;260:404–413. doi: 10.1016/s0012-1606(03)00256-2. [DOI] [PubMed] [Google Scholar]
- 16.Skottman H, Mikkola M, Lundin K, et al. Gene expression signatures of seven individual human embryonic stem cell lines. Stem Cells Oct. 2005;23:1343–1356. doi: 10.1634/stemcells.2004-0341. [DOI] [PubMed] [Google Scholar]
- 17.Ge X, Yamamoto S, Tsutsumi S, et al. Interpreting expression profiles of cancers by genome-wide survey of breadth of expression in normal tissues. Genomics Aug. 2005;86:127–141. doi: 10.1016/j.ygeno.2005.04.008. [DOI] [PubMed] [Google Scholar]
- 18.Lenburg ME, Liou LS, Gerry NP, Frampton GM, Cohen HT, Christman MF. Previously unidentified changes in renal cell carcinoma gene expression identified by parametric analysis of microarray data. BMC Cancer. 2003 Nov 27;3:31. doi: 10.1186/1471-2407-3-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hirst CE, Ng ES, Azzola L, et al. Transcriptional profiling of mouse and human ES cells identifies SLAIN1, a novel stem cell gene. Dev Biol. 2006 May 1;293:90–103. doi: 10.1016/j.ydbio.2006.01.023. [DOI] [PubMed] [Google Scholar]
- 20.Perez-Iratxeta C, Palidwor G, Porter CJ, et al. Study of stem cell function using microarray experiments. FEBS Lett. 2005 Mar 21;579:1795–1801. doi: 10.1016/j.febslet.2005.02.020. [DOI] [PubMed] [Google Scholar]
- 21.Haferlach T, kohlmann A, Basso G, et al. A multi-center and multi-national program to assess the clinical accuracy of the molecular subclassification of leukemia by gene expression profiling. Blood. 2005;106:224a. [Google Scholar]
- 22.Irizarry RA, Warren D, Spencer F, et al. Multiple-laboratory comparison of microarray platforms. Nat Methods May. 2005;2:345–350. doi: 10.1038/nmeth756. [DOI] [PubMed] [Google Scholar]
- 23.Eisen MB, Spellman PT, Brown PO, Botstein D. Cluster analysis and display of genome-wide expression patterns. Proc Natl Acad Sci U S A. 1998;95:14863–14868. doi: 10.1073/pnas.95.25.14863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cowan CA, Klimanskaya I, McMahon J, et al. Derivation of embryonic stem-cell lines from human blastocysts. N Engl J Med. 2004 Mar 25;350:1353–1356. doi: 10.1056/NEJMsr040330. [DOI] [PubMed] [Google Scholar]
- 25.Inzunza J, Gertow K, Stromberg MA, et al. Derivation of human embryonic stem cell lines in serum replacement medium using postnatal human fibroblasts as feeder cells. Stem Cells Apr. 2005;23:544–549. doi: 10.1634/stemcells.2004-0201. [DOI] [PubMed] [Google Scholar]
- 26.Elghetany MT, Patel J. Assessment of CD24 expression on bone marrow neutrophilic granulocytes: CD24 is a marker for the myelocytic stage of development. Am J Hematol Dec. 2002;71:348–349. doi: 10.1002/ajh.10176. [DOI] [PubMed] [Google Scholar]
- 27.Semaphorin-Nomenclature-Committee. Unified nomenclature for the semaphorins/collapsins. Semaphorin Nomenclature Committee. Cell. 1999 May 28;97:551–552. doi: 10.1016/s0092-8674(00)80766-7. [DOI] [PubMed] [Google Scholar]
- 28.Assou S, Anahory T, Pantesco V, et al. The human cumulus-oocyte complex gene expression profile. Human Reproduction. 2006;21:1705. doi: 10.1093/humrep/del065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sperger JM, Chen X, Draper JS, et al. Gene expression patterns in human embryonic stem cells and human pluripotent germ cell tumors. Proc Natl Acad Sci U S A. 2003 Nov 11;100:13350–13355. doi: 10.1073/pnas.2235735100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Brandenberger R, Wei H, Zhang S, et al. Transcriptome characterization elucidates signaling networks that control human ES cell growth and differentiation. Nat Biotechnol Jul. 2004;22:707–716. doi: 10.1038/nbt971. [DOI] [PubMed] [Google Scholar]
- 31.Wei CL, Miura T, Robson P, et al. Transcriptome Profiling of Human and Murine ESCs Identifies Divergent Paths Required to Maintain the Stem Cell State. Stem Cells. 2005;23:166–185. doi: 10.1634/stemcells.2004-0162. [DOI] [PubMed] [Google Scholar]
- 32.Fortunel NO, Otu HH, Ng HH, et al. Comment on “ ‘Stemness’: transcriptional profiling of embryonic and adult stem cells” and “a stem cell molecular signature”. Science. 2003 Oct 17;302:393. doi: 10.1126/science.1086384. author reply 393. [DOI] [PubMed] [Google Scholar]
- 33.Suarez-Farinas M, Noggle S, Heke M, Hemmati-Brivanlou A, Magnasco MO. Comparing independent microarray studies: the case of human embryonic stem cells. BMC Genomics. 2005;6:99. doi: 10.1186/1471-2164-6-99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Johnson J, Bagley J, Skaznik-Wikiel M, et al. Oocyte generation in adult mammalian ovaries by putative germ cells in bone marrow and peripheral blood. Cell. 2005 Jul 29;122:303–315. doi: 10.1016/j.cell.2005.06.031. [DOI] [PubMed] [Google Scholar]
- 35.Cauffman G, Van de Velde H, Liebaers I, Van Steirteghem A. Oct-4 mRNA and protein expression during human preimplantation development. Mol Hum Reprod Mar. 2005;11:173–181. doi: 10.1093/molehr/gah155. [DOI] [PubMed] [Google Scholar]
- 36.Kristiansen G, Schluns K, Yongwei Y, Denkert C, Dietel M, Petersen I. CD24 is an independent prognostic marker of survival in nonsmall cell lung cancer patients. Br J Cancer. 2003 Jan 27;88:231–236. doi: 10.1038/sj.bjc.6600702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lavabre-Bertrand T, Duperray C, Brunet C, et al. Quantification of CD24 and CD45 antigens in parallel allows a precise determination of B-cell maturation stages: relevance for the study of B-cell neoplasias. Leukemia Mar. 1994;8:402–408. [PubMed] [Google Scholar]
- 38.Aigner S, Sthoeger ZM, Fogel M, et al. CD24, a mucin-type glycoprotein, is a ligand for P-selectin on human tumor cells. Blood. 1997 May 1;89:3385–3395. [PubMed] [Google Scholar]
- 39.White JA, Beckett-Jones B, Guo YD, et al. cDNA cloning of human retinoic acid-metabolizing enzyme (hP450RAI) identifies a novel family of cytochromes P450. J Biol Chem. 1997 Jul 25;272:18538–18541. doi: 10.1074/jbc.272.30.18538. [DOI] [PubMed] [Google Scholar]
- 40.Geiman TM, Durum SK, Muegge K. Characterization of gene expression, genomic structure, and chromosomal localization of Hells (Lsh) Genomics. 1998 Dec 15;54:477–483. doi: 10.1006/geno.1998.5557. [DOI] [PubMed] [Google Scholar]
- 41.Sato N, Meijer L, Skaltsounis L, Greengard P, Brivanlou AH. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat Med Jan. 2004;10:55–63. doi: 10.1038/nm979. [DOI] [PubMed] [Google Scholar]
- 42.Ball C, Sherlock G, Parkinson H, et al. A guide to microarray experiments--an open letter to the scientific journals. Lancet. 2002 Sep 28;360:1019. [PubMed] [Google Scholar]
- 43.Abeyta MJ, Clark AT, Rodriguez RT, Bodnar MS, Pera RA, Firpo MT. Unique gene expression signatures of independently-derived human embryonic stem cell lines. Hum Mol Genet. 2004 Mar 15;13:601–608. doi: 10.1093/hmg/ddh068. [DOI] [PubMed] [Google Scholar]
- 44.Aghajanova L, Skottman H, Stromberg AM, Inzunza J, Lahesmaa R, Hovatta O. Expression of leukemia inhibitory factor and its receptors is increased during differentiation of human embryonic stem cells. Fertil Steril Oct. 2006;86 (Suppl 4):1193–1209. doi: 10.1016/j.fertnstert.2005.12.081. [DOI] [PubMed] [Google Scholar]
- 45.Armstrong L, Hughes O, Yung S, et al. The role of PI3K/AKT, MAPK/ERK and NFkappabeta signalling in the maintenance of human embryonic stem cell pluripotency and viability highlighted by transcriptional profiling and functional analysis. Hum Mol Genet. 2006 Jun 1;15:1894–1913. doi: 10.1093/hmg/ddl112. [DOI] [PubMed] [Google Scholar]
- 46.Beqqali A, Kloots J, Ward-van Oostwaard D, Mummery C, Passier R. Genome-wide transcriptional profiling of human embryonic stem cells differentiating to cardiomyocytes. Stem Cells Aug. 2006;24:1956–1967. doi: 10.1634/stemcells.2006-0054. [DOI] [PubMed] [Google Scholar]
- 47.Bhattacharya B, Miura T, Brandenberger R, et al. Gene expression in human embryonic stem cell lines: unique molecular signature. Blood. 2004 Apr 15;103:2956–2964. doi: 10.1182/blood-2003-09-3314. [DOI] [PubMed] [Google Scholar]
- 48.Bhattacharya B, Cai J, Luo Y, et al. Comparison of the gene expression profile of undifferentiated human embryonic stem cell lines and differentiating embryoid bodies. BMC Dev Biol. 2005 Oct 5;5:22. doi: 10.1186/1471-213X-5-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Brandenberger R, Khrebtukova I, Thies RS, et al. MPSS profiling of human embryonic stem cells. BMC Dev Biol. 2004 Sep 10;4:10. doi: 10.1186/1471-213X-4-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Brimble SN, Zeng X, Weiler DA, et al. Karyotypic stability, genotyping, differentiation, feeder-free maintenance, and gene expression sampling in three human embryonic stem cell lines derived prior to August 9, 2001. Stem Cells Dev Dec. 2004;13:585–597. doi: 10.1089/scd.2004.13.585. [DOI] [PubMed] [Google Scholar]
- 51.Brimble SN, Sherrer ES, Uhl EW, et al. The Cell Surface Glycosphingolipids SSEA-3 and SSEA-4 are not Essential for Human ES Cell Pluripotency. Stem Cells. 2006 Sep 28; doi: 10.1634/stemcells.2006-0232. [DOI] [PubMed] [Google Scholar]
- 52.Cai J, Chen J, Liu Y, et al. Assessing self-renewal and differentiation in hESC lines. Stem Cells. 2005 Nov 17; [Google Scholar]
- 53.Calhoun JD, Rao RR, Warrenfeltz S, et al. Transcriptional profiling of initial differentiation events in human embryonic stem cells. Biochem Biophys Res Commun. 2004 Oct 15;323:453–464. doi: 10.1016/j.bbrc.2004.08.117. [DOI] [PubMed] [Google Scholar]
- 54.Darr H, Mayshar Y, Benvenisty N. Overexpression of NANOG in human ES cells enables feeder-free growth while inducing primitive ectoderm features. Development Mar. 2006;133:1193–1201. doi: 10.1242/dev.02286. [DOI] [PubMed] [Google Scholar]
- 55.Dvash T, Mayshar Y, Darr H, et al. Temporal gene expression during differentiation of human embryonic stem cells and embryoid bodies. Hum Reprod Dec. 2004;19:2875–2883. doi: 10.1093/humrep/deh529. [DOI] [PubMed] [Google Scholar]
- 56.Enver T, Soneji S, Joshi C, et al. Cellular differentiation hierarchies in normal and culture-adapted human embryonic stem cells. Hum Mol Genet. 2005 Nov 1;14:3129–3140. doi: 10.1093/hmg/ddi345. [DOI] [PubMed] [Google Scholar]
- 57.Ginis I, Luo Y, Miura T, et al. Differences between human and mouse embryonic stem cells. Dev Biol. 2004 May 15;269:360–380. doi: 10.1016/j.ydbio.2003.12.034. [DOI] [PubMed] [Google Scholar]
- 58.Josephson R, Sykes G, Liu Y, et al. A molecular scheme for improved characterization of human embryonic stem cell lines. BMC Biol. 2006;4:28. doi: 10.1186/1741-7007-4-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kim CG, Lee JJ, Jung DY, et al. Profiling of differentially expressed genes in human stem cells by cDNA microarray. Mol Cells. 2006 Jun 30;21:343–355. [PubMed] [Google Scholar]
- 60.Li H, Liu Y, Shin S, et al. Transcriptome coexpression map of human embryonic stem cells. BMC Genomics. 2006;7:103. doi: 10.1186/1471-2164-7-103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Liu Y, Shin S, Zeng X, et al. Genome wide profiling of human embryonic stem cells (hESCs), their derivatives and embryonal carcinoma cells to develop base profiles of U.S. Federal government approved hESC lines. BMC Dev Biol. 2006;6:20. doi: 10.1186/1471-213X-6-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Miura T, Luo Y, Khrebtukova I, et al. Monitoring early differentiation events in human embryonic stem cells by massively parallel signature sequencing and expressed sequence tag scan. Stem Cells Dev Dec. 2004;13:694–715. doi: 10.1089/scd.2004.13.694. [DOI] [PubMed] [Google Scholar]
- 63.Player A, Wang Y, Bhattacharya B, Rao M, Puri RK, Kawasaki ES. Comparisons between transcriptional regulation and RNA expression in human embryonic stem cell lines. Stem Cells Dev Jun. 2006;15:315–323. doi: 10.1089/scd.2006.15.315. [DOI] [PubMed] [Google Scholar]
- 64.Pyle AD, Donovan PJ, Lock LF. Chipping away at ‘stemness’. Genome Biol. 2004;5:235. doi: 10.1186/gb-2004-5-8-235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Rao M. Conserved and divergent paths that regulate self-renewal in mouse and human embryonic stem cells. Dev Biol. 2004 Nov 15;275:269–286. doi: 10.1016/j.ydbio.2004.08.013. [DOI] [PubMed] [Google Scholar]
- 66.Rao RR, Calhoun JD, Qin X, Rekaya R, Clark JK, Stice SL. Comparative transcriptional profiling of two human embryonic stem cell lines. Biotechnol Bioeng. 2004 Nov 5;88:273–286. doi: 10.1002/bit.20245. [DOI] [PubMed] [Google Scholar]
- 67.Rao RR, Stice SL. Gene expression profiling of embryonic stem cells leads to greater understanding of pluripotency and early developmental events. Biol Reprod Dec. 2004;71:1772–1778. doi: 10.1095/biolreprod.104.030395. [DOI] [PubMed] [Google Scholar]
- 68.Richards M, Tan SP, Tan JH, Chan WK, Bongso A. The transcriptome profile of human embryonic stem cells as defined by SAGE. Stem Cells. 2004;22:51–64. doi: 10.1634/stemcells.22-1-51. [DOI] [PubMed] [Google Scholar]
- 69.Richards M, Tan SP, Chan WK, Bongso A. Reverse serial analysis of gene expression (SAGE) characterization of orphan SAGE tags from human embryonic stem cells identifies the presence of novel transcripts and antisense transcription of key pluripotency genes. Stem Cells May. 2006;24:1162–1173. doi: 10.1634/stemcells.2005-0304. [DOI] [PubMed] [Google Scholar]
- 70.Robson P. The maturing of the human embryonic stem cell transcriptome profile. Trends Biotechnol Dec. 2004;22:609–612. doi: 10.1016/j.tibtech.2004.10.001. [DOI] [PubMed] [Google Scholar]
- 71.Sato N, Brivanlou AH. Microarray approach to identify the signaling network responsible for self-renewal of human embryonic stem cells. Methods Mol Biol. 2006;331:267–283. doi: 10.1385/1-59745-046-4:267. [DOI] [PubMed] [Google Scholar]
- 72.Skottman H, Stromberg AM, Matilainen E, Inzunza J, Hovatta O, Lahesmaa R. Unique gene expression signature by human embryonic stem cells cultured under serum-free conditions correlates with their enhanced and prolonged growth in an undifferentiated stage. Stem Cells Jan. 2006;24:151–167. doi: 10.1634/stemcells.2004-0189. [DOI] [PubMed] [Google Scholar]
- 73.Sun BW, Yang AC, Feng Y, et al. Temporal and parental-specific expression of imprinted genes in a newly derived Chinese human embryonic stem cell line and embryoid bodies. Hum Mol Genet. 2006 Jan 1;15:65–75. doi: 10.1093/hmg/ddi427. [DOI] [PubMed] [Google Scholar]
- 74.Xu RH, Chen X, Li DS, et al. BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nat Biotechnol Dec. 2002;20:1261–1264. doi: 10.1038/nbt761. [DOI] [PubMed] [Google Scholar]
- 75.Zeng X, Miura T, Luo Y, et al. Properties of pluripotent human embryonic stem cells BG01 and BG02. Stem Cells. 2004;22:292–312. doi: 10.1634/stemcells.22-3-292. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.