

Abstract
The survival of motor neuron (SMN) protein is mutated in patients with spinal muscular atrophy (SMA). SMN is part of a multiprotein complex required for biogenesis of the Sm class of small nuclear ribonucleoproteins (snRNPs). Following assembly of the Sm core domain, snRNPs are transported to the nucleus via importin β. Sm snRNPs contain a nuclear localization signal (NLS) consisting of a 2,2,7-trimethylguanosine (TMG) cap and the Sm core. Snurportin1 (SPN) is the adaptor protein that recognizes both the TMG cap and importin β. Here, we report that a mutant SPN construct lacking the importin β binding domain (IBB), but containing an intact TMG cap-binding domain, localizes primarily to the nucleus, whereas full-length SPN localizes to the cytoplasm. The nuclear localization of the mutant SPN was not a result of passive diffusion through the nuclear pores. Importantly, we found that SPN interacts with SMN, Gemin3, Sm snRNPs and importin β. In the presence of ribonucleases, the interactions with SMN and Sm proteins were abolished, indicating that snRNAs mediate this interplay. Cell fractionation studies showed that SPN binds preferentially to cytoplasmic SMN complexes. Notably, we found that SMN directly interacts with importin β in a GST-pulldown assay, suggesting that the SMN complex might represent the Sm core NLS receptor predicted by previous studies. Therefore, we conclude that, following Sm protein assembly, the SMN complex persists until the final stages of cytoplasmic snRNP maturation and may provide somatic cell RNPs with an alternative NLS.
INTRODUCTION
The nuclear periphery is a very busy place: more than a million macromolecules are thought to be actively transported between the nucleus and the cytoplasm each minute (1-4). This bidirectional traffic is routed through nuclear pore complexes (NPCs), large protein structures that span the nuclear membrane bilayer (2,5,6). The NPCs are active participants in nucleocytoplasmic transport, which is mediated by soluble transport receptors. These receptor proteins, collectively known as karyopherins, shuttle between the nucleus and cytoplasm, carrying a wide variety of cargoes. Unidirectional transport of the cargo molecules is achieved by two families of karyopherins called importins and exportins (7). Directionality is imposed on nucleocytoplasmic traffic by a small GTPase called Ran. The concentration of RanGTP is higher in the nucleus than in the cytoplasm, and this gradient is maintained by the restricted subcellular localization of the guanine nucleotide exchange factor (RCC1), which is anchored to the chromatin (8,9). Cargoes identify themselves to the nucleocytoplasmic transport machinery by motifs called nuclear localization signals (NLSs) and nuclear export signals (NESs). These signals can be protein- or RNA-based or a composite of the two (10-14). Many classes of cargo contain signals that bind directly to a cognate receptor, while others have signals that bind to the receptor via adaptor proteins.
Small nuclear RNAs (snRNAs) of the Sm class have a rather unique life-cycle (15). Following transcription in the nucleus, these RNA pol II transcripts are exported to the cytoplasm by binding to an adaptor protein called PHAX (16). Assembly into stable ribonucleoprotein particles (snRNPs) requires the activity of the survival of motor neurons (SMN) protein complex (reviewed in 15 and 17). After assembly of the heteroheptameric Sm-ring, the 5′-end methylguanosine cap structure is hypermethylated (18) to form a trimethylguanosine (TMG) cap by a protein called Tgs1 (19,20). Cap hypermethylation is a signal that triggers nuclear import (11,21-23) via binding to a protein called snurportin1 (24). Snurportin1 (SPN) is thus an adaptor that links snRNP cargo to importin β for subsequent transport to the nucleus (24). The N-terminal, importin β binding (IBB) domain of SPN (Fig. 1A) shares significant similarity with importin α family members (24). However, unlike other importins, SPN contains neither an NLS-binding domain, nor a domain that recognizes RanGTP (24,25). In contrast to that of importin a, the IBB domain of SPN allows for snRNP cargo to be imported in a Ran- and energy-independent fashion (26). It is important to note that SPN binds only the TMG cap and not to the Sm core (24). Thus, the C-terminus of SPN contains a novel TMG-binding domain.
Figure 1.
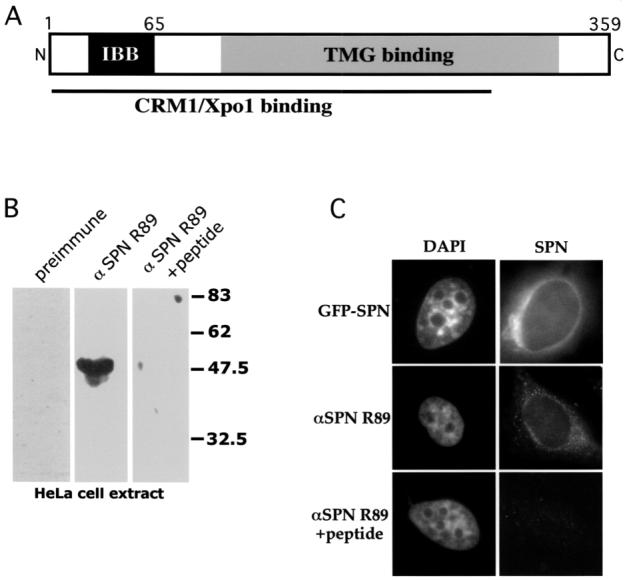
Schematic of snurportin1 (SPN) and characterization of antibody R89. (A) A cartoon of SPN, indicating the importin β (IBB), the CRM1/Xpo1 and trimethylguanosine (TMG) cap binding domains (24). (B) Western blot analysis of HeLa extract with SPN antibody R89. The left lane was probed with preimmune serum, the middle lane with R89 and the right lane with R89 in the presence of a 10-fold excess of the cognate peptide. The antibody specifically detected a band of approximately 45 kDa that was competed by the peptide. (C) The subcellular localization of SPN was studied by transient transfection of HeLa cells with GFP-SPN (top panels). In the middle panels, cells were immunostained with anti-SPN R89. Competition with a 20-fold excess of peptide (lower panels) resulted in background levels of fluorescence. Both the exogenous and endogenous SPN localized to the cytoplasm with enrichment at the nuclear periphery.
Microinjection studies in Xenopus laevis oocytes have shown that Sm snRNPs (U1, U2, U4 and U5) possess a complex NLS composed of both the Sm core domain and the TMG cap (22,23,27). Interestingly, not all the spliceosomal snRNAs display the same TMG cap requirements for import in oocytes. Whereas U1 and U2 snRNA absolutely require an intact cap, U4 and U5 snRNAs can be imported as ApppG-capped derivatives, albeit with reduced transport kinetics (23). In contrast, the TMG cap is not essential for snRNP import in somatic cells (28), although it accelerates the rate of transport. The Sm core domain is therefore necessary and sufficient to mediate nuclear targeting of snRNPs in somatic cells.
We were interested in perturbing the rate of Sm snRNP import in order to assay the effect on a prominent nuclear subdomain called the Cajal body (reviewed in 29 and 30). Cajal bodies (CBs) are the first detectable sites of nuclear accumulation of newly imported Sm snRNPs (31). Furthermore, CBs associate with specific chromosomal loci in interphase cells (32-34); colocalization with the U2 snRNA gene cluster requires not only transcriptionally competent U2 loci (35) but the presence of intact U2 snRNPs (36). Since Huber et al. (24) showed that an N-terminal deletion of SPN (SPNΔN65) was dominant negative for snRNP import in Xenopus oocytes, we decided to test the efficacy of this mutant in the mammalian system. To our surprise, SPNΔN65 overexpression did not allow us to assay the interaction of CBs and snRNA genes, since it failed to titrate newly assembled snRNPs in the cytoplasm. Instead, we found that SPNΔN65 was actively imported into the nucleus, consistent with the existence of a parallel, cap-independent snRNP import pathway in somatic cells (11,28). Furthermore, SPN and SPNΔN65 overexpression induced disassembly of CBs and SMN gems.
Previous studies suggested that a cytoplasmic factor bound to the Sm core domain mediates snRNP import in the absence of a functional TMG cap receptor (11,23,37). We hypothesized that the SMN complex, which is required for assembly of the Sm ring (15,38 and references therein), might provide this alternative import signal. Significantly, we found that SPN and SMN interact in GST-pulldown and coimmunoprecipitation assays. The interaction of SPN and SMN is not direct, but mediated by RNA, and is restricted to the cytoplasmic compartment. Gel filtration assays confirmed that SMN, SPN and importin β cofractionate. As expected, only a relatively small fraction of the total SMN protein comigrates with importin β. Intriguingly, a zinc finger protein called ZPR1, implicated in the nuclear targeting of SMN (39), was also found to interact with a complex that contains SPN. These results demonstrate that SMN is not only present during early cytoplasmic steps of Sm assembly but also remains in the snRNP complex through late steps. Consistent with the idea that SMN might accompany newly assembled snRNPs into the nucleus, we also found that recombinant SMN interacts directly with importin β. Taken together, these results establish the existence of an import-competent snRNP complex that contains SMN and importin β.
RESULTS
Snurportin1 is primarily a cytoplasmic protein
Snurportin1 is a protein of approximately 45 kDa that is required for nuclear snRNP import in Xenopus oocytes (24). In order to better characterize the complexes that carry out snRNP import in mammals, we developed polyclonal anti-peptide antibodies against human SPN and cloned the cDNA by RT-PCR using the published sequence (24). Western blot analysis with anti-SPN serum R89 identified a single band of the expected size that was specifically competed upon preincubation with the antigenic peptide (Fig. 1B). Immunofluorescence with this antiserum revealed that the protein is localized throughout the cytoplasm, concentrating near the nuclear periphery (Fig. 1C). This localization pattern is completely consistent with snurportin’s role as a nuclear import adaptor. Equivalent results were obtained upon expression of GFP-SPN (Fig. 1C). Furthermore, incubation of the antibody with excess peptide antigen completely blocked the immunofluorescence signal (Fig. 1C). Western analysis of extracts from cells transfected with GFP-SPN revealed the presence of an additional band of the appropriate size (data not shown). Thus SPN antiserum R89 specifically recognizes both the exogenous and endogenous snurportins.
A snurportin mutant is actively transported to the nucleus
Huber et al. (24) showed that the N-terminal 65 amino acids of snurportin1 contain a domain (IBB, Fig. 1A) required for binding to importin β. Overexpression of SPNΔN65 in frog oocytes abolished import of snRNPs into the nucleus (24). Thus we were quite surprised to find that GFP-SPNΔN65 was primarily nuclear when expressed in human cells (Fig. 2). The molecular mass cutoff for proteins passively diffusing through the NPC is reportedly ∼40-60 kDa (2,40) and depends to some extent on the shape of the macromolecule. That is, proteins larger than this require assistance from importins and exportins. GFP-SPNΔN65 contains an intact TMG-binding domain but cannot bind to importin β (Fig. 1A). Therefore, it was formally possible that the construct could simply diffuse into the nucleus and bind to snRNPs. In the case of wild-type SPN, cargo release is effected upon binding to the export factor CRM1/Xpo1 (25). However, binding to CRM1/Xpo1 and recycling to the cytoplasm is disrupted in SPNΔN65, as export depends on multiple sites throughout the length of the protein (25). Only the C-terminal 75 amino acids are dispensable for this activity (Fig. 1A). Since it was conceivable that GFP-SPNΔN65 (calc. MW ∼65 kDa) was small enough to pass through the nuclear pores unassisted, we cloned a second GFP domain in-frame with GFP-SPN and GFP-SPNΔN65 to ensure that these constructs exceeded the limit for passive diffusion. As shown in Figure 2A, the localization patterns of the 2 ×GFP constructs were identical to those of their 1 × GFP counterparts. These results indicate that GFP-SPNΔN65 is actively imported into the nucleus by an alternative pathway that does not involve direct binding to importin β (11,23,24,37).
Figure 2.
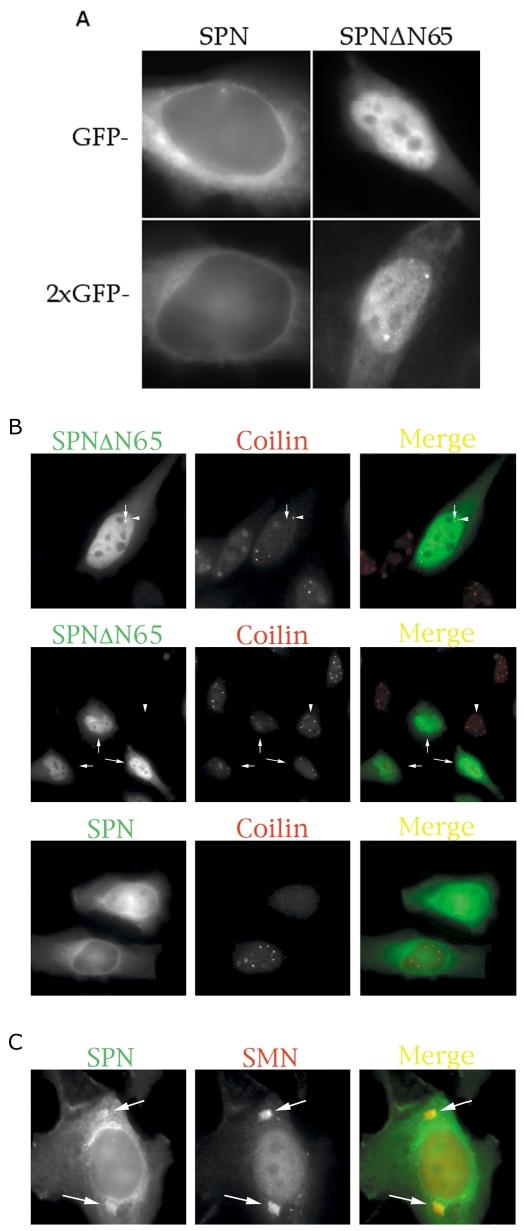
A) SPN is actively imported into the nucleus. Transient transfections of HeLa cells with 1 × or 2 × GFP tags. SPN (left panels) and SPNΔN65 (right) are shown. Surprisingly, SPNΔN65, which lacks an IBB domain, localizes primarily to the nucleus. (B) Comparison of mutant and wild-type proteins and their effects on Cajal bodies (CBs). Transient overexpression of GFP-SPN or GFP-SPNΔN65 causes a reduction in the number of CBs (see Table 1). The larger foci that were occasionally observed upon transfection with GFP-SPNΔN65 (top panels, arrow) did not strictly correspond to CBs (arrowhead). The low-magnification image in the second set of panels shows that cells expressing GFP-SPNΔN65 displayed fewer CBs (arrows) than did the untransfected cells (e.g. the cell marked by the arrowhead). Cells expressing lower levels of GFP-SPN (third panel, lower cell) typically displayed higher numbers of CBs than did those cells expressing higher levels of the construct (upper cell). (C) Cytoplasmic accumulations of SMN (arrows) could sometimes be detected in the GFP-SPN channel (arrows) and were not visible in the DAPI channel (not shown).
Overexpression of SPN and SPNΔN65 disassembles Cajal bodies
In the course of our localization experiments, we noticed that GFP-SPNΔN65 occasionally displayed nuclear foci (Fig. 2A). Immunofluorescence with CB marker proteins coilin and SMN revealed that these focal accumulations did not strictly correspond to CBs, although they sometimes colocalized (Fig. 2B and data not shown). Strikingly, we found that cells transfected with GFP-SPNΔN65 typically displayed fewer CBs than did neighboring untransfected cells (Fig. 2B). In fact, careful analysis revealed that even cells transfected with wild-type GFP-SPN showed a reduction in CB number (Table 1). Whereas moderate levels of GFP-SPNΔN65 expression typically resulted in CB disassembly, only high levels of GFP-SPN had this effect (Fig. 2B). Similar results were obtained in parallel experiments when SMN was used as the CB marker (Table 1). This effect was specific to SPN, as cells transfected with GFP alone showed no difference in CB numbers when compared to untransfected cells (Table 1).
Table 1.
Effects of SPN overexpression on CBs
| Number of foci Coilin | SMN | ||
|---|---|---|---|
| GFP-SPN | Mean | 3.6±2.0 | 3.2±1.7 |
| p-value | 2.2±10-7 | 4.4±10-8 | |
| GFP-SPNΔN65 | Mean | 2.5±2.0 | 1.6±1.6 |
| p-value | 6.0±10-18 | 3.7±10-22 | |
| GFP | Mean | 5.1±1.9 | 5.1±2.7 |
| p-value | NS* | NS* | |
| Untransfected | 5.4±2.0 | 5.2±2.1 |
NS, not significant.
As mentioned above, the typical SPN localization pattern was diffuse throughout the cytoplasm, with a relatively pronounced staining of the nuclear periphery. Thus the majority of cells did not show any obvious colocalization with the cytoplasmic fraction of SMN (data not shown). However, we sometimes observed local accumulations of both GFP-SPN and SMN in the cytoplasm (Fig. 2C), suggesting a possible interaction between the snRNP assembly and import machineries.
Snurportin1 forms a cytoplasmic complex with SMN and splicing snRNPs
The SMN complex is required for assembly of the Sm ring which, in turn, is required for cap hypermethylation (reviewed in 15). Precisely when SMN leaves the snRNP complex following Sm core assembly has not been established. In order to determine whether SMN and SPN are capable of binding to the same RNPs, we performed GST-pulldown assays with HeLa cytoplasmic extracts. As shown in Figure 3A, SMN, Gemin3 and SmB/B’ proteins were recovered by GST-SPN bound to glutathione-sepharose beads. Control experiments with GST alone showed only residual binding. Thus the SMN complex remains bound to TMG-positive Sm snRNPs. In order to prove that SPN forms a native complex with SMN, cells were transfected with GFP-tagged SPN constructs and analyzed by co-immunoprecipitation with anti-GFP antibodies. As shown in Figure 3B, SMN was coprecipitated by both GFP-SPN and GFP-SPNΔN65. Parallel northern blotting experiments (Figs 3C and D) showed that U2 snRNA was also coprecipitated with the SPN constructs. Both wild-type SPN and the IBB mutant protein have intact TMG-binding domains and, as expected, each bound equivalent amounts of U2 snRNA in the GST-pulldown experiment (Fig. 3C). However, quantification of the blot in Figure 3D showed that GFP-SPNΔN65 brought down roughly five times more U2 RNA than did GFP-SPN and 10 times more than GFP alone. GFP-SPN brought down twice as much RNA as did the GFP control. Given the nuclear localization of both GFP-SPNΔN65 and the bulk of the Sm snRNPs, this result is not surprising. Additionally, SPN binding to CRM1/Xpo1 is required for cargo release and cytoplasmic recycling. Thus the SPNΔN65 construct, which cannot bind to CRM1, likely binds ‘irreversibly’ to snRNPs, whereas the wild-type protein does not.
Figure 3.
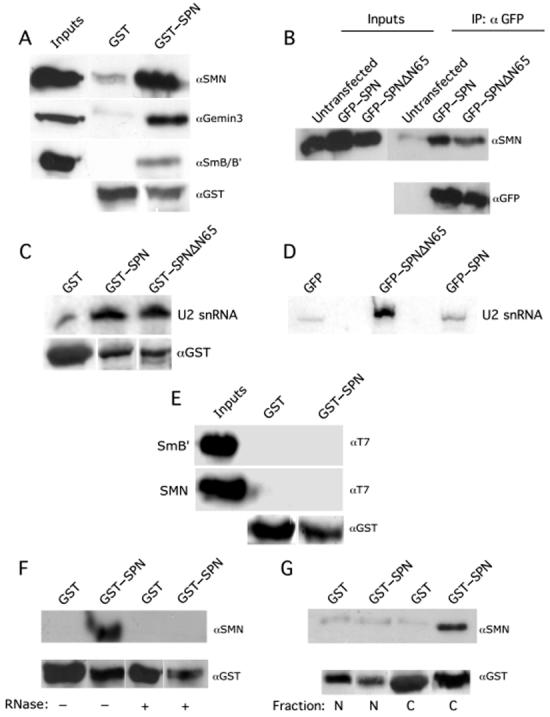
SPN interacts indirectly with SMN in the cytoplasm. (A) SPN interacts with the cytoplasmic SMN complex in vitro. Pulldown assays with GST-SPN or GST alone were performed using HeLa cytoplasmic extracts. The pulldowns were analyzed by western blotting with monoclonal antibodies against SMN (7B10), Gemin3 (11G9) and SmB/B′(Y12). (B) SPN interacts with SMN in vivo. HeLa cells were transiently transfected with GFP-SPN or GFP-SPNΔN65; untransfected cells were used as a negative control. Immunoprecipitations (IPs) were performed from total HeLa cell lysates with polyclonal antibodies against GFP. The IPs were then analyzed by western blotting with anti-SMN. (C) SPN and SPNΔN65 interact with U2 snRNA with similar affinities in vitro. GST-pulldown assays were performed using GST-SPN, GST-SPNΔN65 and GST (negative control) from total Hela cell lysate. RNA was isolated from the pulldowns; a U2 snRNA specific radiolabeled probe was used for analysis by northern blotting. The lower panel shows the loading controls; note that the GST-only lane is slightly overloaded relative to the experimental lanes. (D) SPNΔN65 and SPN bind U2 snRNA in vivo. HeLa cells were transiently transfected with GFP-SPN, GFP-SPNΔN65 or GFP (negative control). IPs were performed from total HeLa cell lysates with monoclonal antibodies against GFP. RNA was isolated from the IP, and analyzed by northern blotting. (E) SPN and SMN do not interact directly. Pulldown assays were performed using GST-SPN and GST (negative control), along with His-T7-SmB′or His-T7-SMN. The pulldowns were analyzed by western blotting with anti-T7. (F) RNA mediates the interaction of SPN with SMN. Pulldown assays were performed using GST-SPN or GST (negative control) from HeLa cell lysates in the absence or presence of ribonucleases. The pulldowns were analyzed by western blotting with anti-SMN. (G) SPN interacts with cytoplasmic SMN. Pulldown assays were performed using GST-SPN and GST (negative control) from HeLa cells (nuclear or cytoplasmic fractions). The pulldowns were analyzed by western blotting with anti-SMN. Anti-GST and anti-GFP were used as loading controls where indicated. Inputs show 12% of the total lysate used in the pulldowns.
To test whether SMN and SPN might interact directly, we purified recombinant proteins expressed in bacteria and performed GST-pulldowns. No direct binding was detected (Fig. 3E). Therefore, the interaction between SPN and SMN is likely mediated by the snRNA. To test this hypothesis, we performed GST-pulldown assays from cell extracts incubated in the presence or absence of RNases A and T1. Figure 3F reveals that SMN was efficiently recovered in the absence of added RNase and not detectable in its presence. The blots were reprobed with anti-GST to demonstrate that roughly equivalent amounts of protein were used. As an independent control for monitoring RNase activity, extracts were incubated in the presence or absence of RNase and then assayed for binding of U2B″, a U2-specific snRNP protein, to U2 RNA (data not shown).
In order to determine if SMN and SPN complexes were restricted to a particular subcellular compartment, nuclear and cytoplasmic HeLa cell extracts were prepared and GST-pulldown assays performed with GST-SPN and GST alone. Cell fractionation was monitored using control antibodies (data not shown) against proteins that localize to the cytoplasm [glutaredoxin, (41)] and nucleus [coilin, (42)]. As shown in Figure 3G, SMN was only recovered from the cytoplasmic fractions. Taken together, these data establish that SPN and SMN co-exist in a cytoplasmic complex, bridged by snRNAs. Thus we conclude that SMN remains bound to Sm snRNPs after cap hypermethylation, forming a putative pre-import complex.
SMN and snurportin form a complex that contains importin β
The interaction of Sm snRNPs, SPN and importin β has been well established (24). Our finding that SMN and SPN form a complex together prompted the question of whether SMN is present in an import-competent snRNP complex with importin β. As a first step, we transfected cells with myc-SMN, immunoprecipitated with anti-myc antibodies, and probed a western blot with anti-importin β. Interestingly, Figure 4A shows that SMN exists in a complex with importin β in vivo. In order to show that SPN, SMN and importin β are together in the same complexes, gel filtration assays were performed using HeLa cytoplasmic extract, followed by western blot analysis. SMN runs in two peaks, the first a broad one between 600 and 1500 kDa and the other between 200 and 500 kDa (Fig. 4C). Notably, importin β, SMN and SPN comigrate in the 400-500 kDa region, suggesting that these proteins form a single complex.
Figure 4.
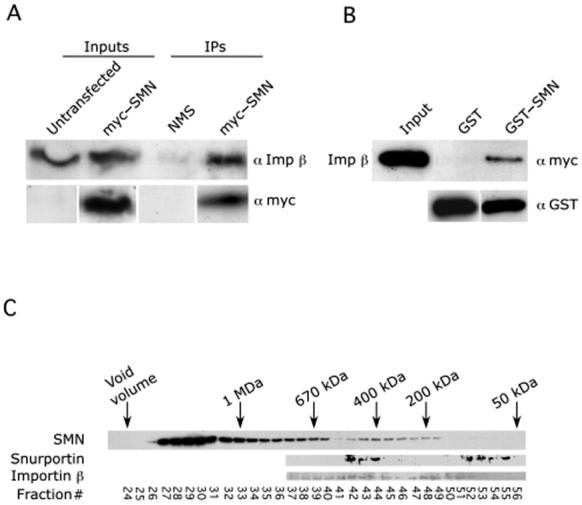
SMN interacts directly with importin β. (A) HeLa cells were transfected with myc-SMN; untransfected cells were used as a negative control. Immunoprecipitations (IPs) were performed from total HeLa cell lysates with monoclonal antibodies against myc. The IPs were then analyzed by western blotting with anti-Imp β. (B) SMN and Imp β interact directly using purified recombinant proteins. Pulldown assays were performed using GST-SMN or GST alone and His-myc-importin β. The pulldowns were analyzed by western blotting with anti-myc and anti-GST (loading control). Input shows approximately 50% of the total lysate used in the pulldowns. (C) Gel filtration column of HeLa cytoplasm. Fractions were analyzed by western blotting with the indicated antibodies. SPN, SMN and importin β cofractionate in the ∼400 kDa range, consistent with the existence of a complex containing all three proteins.
As described above, several lines of evidence suggest the existence of an Sm core-directed, TMG cap-independent import mechanism that also impinges upon the importin β pathway (11 and references therein). In light of our findings, the simplest explanation for these data is that the SMN complex is the factor that recognizes both the Sm core and importin β, thus acting as the adaptor for this cap-independent pathway. We decided to test this idea further by asking whether SMN and importin β could bind to each other directly using recombinant proteins. As shown in Figure 4B, GST-SMN, but not GST alone, clearly interacted with His-tagged importin β.
Snurportin, SMN and the zinc finger protein ZPR1 form a complex
Given that SPN, importin β and SMN form what appears to be an import-competent complex, this raised the question as to whether the SMN complex is imported into the nucleus along with the newly assembled snRNPs. A recent study found that the zinc finger protein ZPR1 is required for the accumulation of SMN in the nucleus (39). ZPR1 is an essential protein in yeast (44). In mammals, the protein is involved in relaying proliferative growth signals from the cell surface to the nucleus (45,46). Although ZPR1 is localized to the cytoplasm in quiescent cells, the protein is translocated to the nucleus upon mitogen stimulation (44-46). Truncation of the C-terminus of ZPR1 abolished the interaction with SMN and was dominant negative for SMN localization to nuclear bodies (39). Therefore, we wanted to determine whether ZPR1 might also be complexed with SPN. Toward that end, we performed GST-pulldowns and co-immunoprecipitations. HeLa cell lysate was passed over GST and GST-SPN beads and then probed with anti-ZPR1 antibodies (Fig. 5A), revealing an interaction between the two proteins. Similarly, lysates from cells transfected with GFP-SPN were immunoprecipitated with anti-GFP antibodies and probed for the presence of ZPR1. As shown in Figure 5B, GFP-SPN but not GFP alone brought down ZPR1 protein. The interaction between SPN and ZPR1 was sensitive to RNase digestion (Fig. 5C) and was primarily restricted to the cytoplasm (Fig. 5D). Furthermore, gel fractionation studies showed that ZPR1 also comigrated with SPN, SMN and importin β (data not shown), suggesting that these factors are present in the same complexes. Thus the fact that ZPR1, which is involved in regulating the accumulation of SMN in the nucleus (39), is also complexed with SPN and importin β suggests that SMN could indeed accompany newly assembled snRNPs across the NPC and perhaps help to target them to CBs (47,48).
Figure 5.
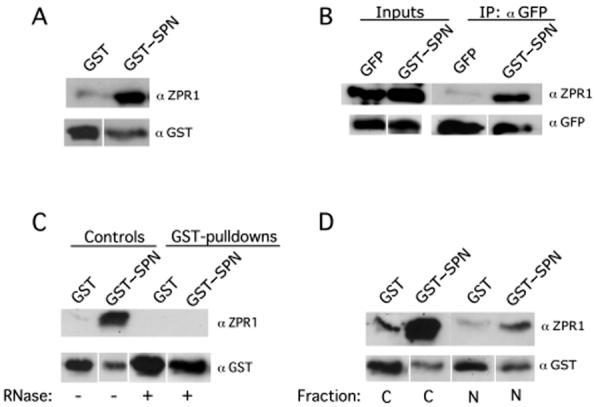
SPN forms a cytoplasmic complex with ZPR1. (A) SPN interacts with ZPR1 in vitro. Pulldown assays with GST-SPN and GST were performed using HeLa lysate. The pulldowns were analyzed by western blotting with monoclonal antibodies against ZPR1 (LG9). (B) SPN interacts with ZPR1 in vivo. HeLa cells were transiently transfected with GFP-SPN or GFP alone. Immunoprecipitations (IPs) were performed from total HeLa cell lysates with polyclonal antibodies against GFP. The IPs were then analyzed by western blotting with LG9 and GFP (loading control). (C) RNA mediates the SPN-ZPR1 interaction. Pulldown assays were performed using GST-SPN; GST (negative control) from HeLa cell lysates in the presence or absence of ribonucleases. (D) SPN interacts with cytoplasmic ZPR1. Pulldown assays were performed using GST-SPN and GST (negative control) along with nuclear (N) or cytoplasmic (C) HeLa cell extracts.
DISCUSSION
One function of the SMN protein complex is to assemble the Sm core domain (38,49). To accomplish this task, members of the complex must contact both the snRNA (50) and the Sm proteins (43,51-53). In vivo, a subset of the Sm proteins are targeted to the SMN complex by symmetrical dimethylarginine residues that are posttranslationally added to their arginine- and glycine-rich tails (54-58). The precise mechanism used by SMN to direct cytoplasmic assembly of the heptameric Sm ring is unknown. Following assembly, the events whereby the newly assembled snRNP particle acquires its hypermethylated 5′ cap and is transported across the nuclear pore are also poorly understood. Our results demonstrate that SMN does not fall off immediately following assembly, but remains associated with the snRNP throughout the downstream cytoplasmic maturation steps. Together with data from the literature, our results suggest the existence of at least three distinct cytoplasmic SMN-snRNP subcomplexes: 1) a post-export, pre-Sm assembly particle (50); 2) one that contains an Sm ring but lacks a TMG cap (59); and 3) a pre-import complex that contains SPN and importin β (this work).
Furthermore, the results suggest a role for the SMN complex in TMG cap formation, since SMN is present both before and after cap hypermethylation. It is therefore possible that the SMN complex also functions to recruit the requisite methyl-transferase. The enzyme that performs this function has recently been identified in both yeast and mammals (19,20). Interestingly, the subcellular distribution of this protein, called Tgs1, mirrors that of SMN: it is diffusely localized throughout the cytoplasm and concentrated in nuclear CBs (20).
Finally, our findings that SMN directly interacts with importin β and that snurportin1, SMN, ZPR1, importin β and TMG-capped snRNPs form a complex indicates that SMN may be imported into the nucleus along with the snRNPs. The simplest explanation to emerge from these data is that the SMN complex is, in fact, the long-awaited Sm core NLS receptor (Fig. 6). Consistent with this interpretation, heterokaryon fusion experiments using FP-tagged Sm proteins showed that only SMN-positive CBs accumulated newly imported proteins (48). Recent work from our laboratory has shown that coilin contains symmetrical dimethylarginine residues (M. D. Hebert and A. G. Matera, unpublished data) and can compete with Sm proteins for binding sites on SMN (47). Thus it is possible that coilin may help to dissociate the SMN-snRNP complex following nuclear import (47). However, direct evidence of SMN crossing the pore along with the snRNPs is currently lacking. Alternatively, SMN may be released from snRNPs just prior to, or concomitant with, import. In that case, nuclear SMN may be involved in other aspects of assembly or reassembly of snRNPs and snoRNPs (60,61), although these two functions need not be mutually exclusive.
Figure 6.
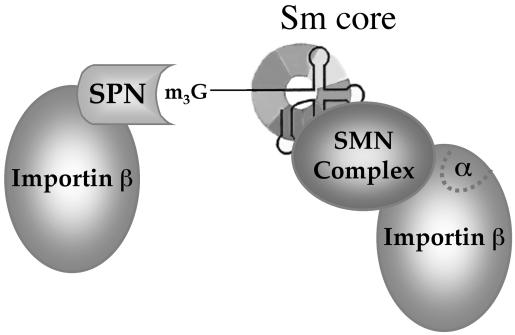
Model of the U snRNP import complex, adapted from (11). SPN binds to the TMG cap (m3G) of Sm class snRNPs and interacts with importin β via its IBB domain. The hypothetical snRNP core-binding factor (see text) is predicted to be the SMN complex. Since SMN lacks an IBB domain, direct binding to importin β (Fig. 4B) is therefore unlikely to proceed via the SPN/importin α binding pocket (α). Two importin β molecules are shown in the model, as predicted by (11). Alternatively, binding of the Sm core by SMN might stabilize the binding of a single importin β. Additional experiments will be required to distinguish between these two possibilities.
In summary, we have shown that SMN is associated with snRNPs during the final steps in cytoplasmic Sm snRNP assembly and that it directly interacts with importin β. Thus the SMN complex is uniquely positioned to act as a quality control factor at multiple steps in the Sm snRNP maturation process. As such, the complex may act as a kind of ‘master assembler’ or chaperone of RNP assembly and transport (62,63). In the future, elucidating the molecular mechanisms behind these processes should help to better distinguish the roles played by SMN in small RNA metabolism and spinal muscular atrophy.
MATERIALS AND METHODS
DNA constructs
Snurportin1 (SPN) and SPNΔN65 were amplified from a PCR-ready human brain cDNA library (Clontech Laboratories, Palo Alto, CA, USA) and cloned into pCR 2.1 (Invitrogen, Carlsbad, CA, USA). Primers containing EcoRI and BamHI restriction sites were used to PCR amplify SPN from pCR 2.1. SPN was subsequently cloned into EcoRI/BamHI-cut pEGFP-C3 (Clontech Laboratories). 2 × EGFP was generated by PCR amplification of EGFP from pEGFP-C3 using primers containing BglII and HindIII restriction sites followed by cloning into BglII/HindIII-digested pEGFP-C3. SPN and SPNΔN65 were then excised from pEGFP-C3-SPN and pEGFP-C3-SPNΔN65, respectively, with EcoRI and BamHI and cloned into EcoRI/BamHI-digested 2 × EGFP-C3, resulting in 2 × EGFP-SPN and 2 × EGFP-SPN1-DN65. Glutathione S-transferase (GST) fusions of SPN and SPNΔN65 were generated by PCR amplifying SPN from 2 × EGFP-C3 SPN with EcoRI forward and BamHI reverse primers. This fragment was subsequently cloned into a BamHI/EcoRI-digested pGex 2T vector (Amersham Pharmacia, Piscataway, NJ, USA).
Antibodies
A polyclonal SPN antibody was generated (Sigma Genosys, The Woodlands, TX, USA) using a synthesized peptide consisting of CGMESEEENKKDDEE residues corresponding to amino acids 73-85 of human SPN. Other antibodies utilized in this study include: mouse monoclonal anti-SMN (7B10; 52); mouse monoclonal anti-ZPR (LG1; 39); mouse monoclonal anti-Gemin 3 (11G9; 43); mouse monoclonal anti-Sm (Y12; 64); rabbit polyclonal anti-importin beta (gift of K. Weis); mouse monoclonal anti-GST (Santa Cruz, CA, USA); rabbit polyclonal and mouse monoclonal anti-GFP (Roche, Indiana-polis, USA); mouse monoclonal anti-myc (9E10; Santa Cruz); mouse monoclonal anti-T7 (Novagen, Madison, WI, USA); rabbit polyclonal anti-coilin (R288; 65); mouse monoclonal anti-U2B″ (gift of H. Salz); and rabbit polyclonal antiglutaredoxin (gift of J. Mieyal). Secondary antibodies utilized were goat anti-mouse and goat anti-rabbit conjugated horseradish peroxidase (Pierce Chemical, Rockford, IL, USA) as well as goat anti-rabbit conjugated fluorescein (Vector Laboratories, Burlingame, CA, USA).
Co-immunoprecipitation
HeLa cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) (Gibco BRL, Gaithersburg, MD, USA) with 10% fetal bovine serum (FBS) (Gibco BRL), penicillin and streptomycin. pEGFP-SPN or pEGFP-SPNΔN65 was transfected into HeLa cells using SuperFect (QIAGEN, Valencia, CA, USA). As a control, untransfected HeLa cells were also utilized. Cells were harvested 24 h after transfection, washed with phosphate-buffered saline (PBS) and resuspended in 1 μl of mRIPA buffer (50 mm Tris-Cl, pH 7.5, 150 mm NaCl, 1% NP-40, 1 mm EDTA) plus protease inhibitor cocktail tablets (Roche) to lyse cells. Resuspended cells were nutated at 4 °C for 30 min, followed by centrifugation for 5 min to pellet cellular debris. Where indicated, 5 mg of total protein from the lysate was treated with RNase A (1 mg, Sigma, St. Louis, MO, USA) and RNase T1 (5000 units, gift of H. Salz) for 1 h at 30 °C. Fifty microliters of polyclonal anti-GFP (Clontech Laboratories) were then added to 900 μl of lysate. After nutating for 1 h, 60 μl of 50% protein A Sepharose beads (Amersham Pharmacia) were added to the lysates, and incubated overnight. Beads were then washed six times with 1 μl of mRIPA, resuspended in 15 μl of 5×sodium dodecyl sulfate (SDS) loading buffer, and boiled, and proteins were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). After transfer to nitrocellulose, membranes were probed with anti-SMN (7B10) followed by incubation with goat anti-mouse conjugated horseradish peroxidase (Pierce Chemical) and chemiluminescence detection (Roche).
Pulldowns
Expression and isolation of recombinant proteins was performed using standard techniques (66). GST pulldowns were conducted by incubating HeLa cell lysate, prepared as described (66), with glutathione bead captured GST tagged proteins for 1 h in a buffer containing 450 mm Na+. Beads were then washed six times with 1 μl of mRIPA, resuspended in 15 μl of 5 × SDS loading buffer, and boiled, and proteins were resolved by SDS-PAGE. Following transfer to nitrocellulose, membranes were probed with appropriate primary and secondary antibodies.
Immunofluorescence
HeLa cells were grown on slides (Nunc, Rochester, NY, USA) to 80% confluency. Slides were fixed in 4% paraformaldehyde for 10 min at room temperature, followed by extraction with 0.5% Triton for 5 min. Incubation in 10% normal goat serum was followed by the addition of anti-SPN1 rabbit polyclonal antibody, R89 and detection with fluorescein-conjugated goat anti-rabbit (Molecular Probes, Eugene, OR, USA). Incubations at 30 °C for 30 min were followed with three washes for 5 min each at room temperature with 1 × PBS.
Protein fractionation
The S100 cytoplasmic fraction was generated from 500 μl of packed HeLa cells (National Cell Culture Center) using the NE-PER extraction system (Pierce Chemical). Five hundred microliters of S100 containing 10.5 mg of total protein were passed over a Superose 6HR 10/30 column and fractionated (300 μl fractions) using fast performance liquid chromatography (FPLC). Twenty-five microliters of each fraction were added to 5 μl of 5 SDS-loading buffer, boiled and subjected to SDS-PAGE. Thirty microliters of S100 fraction were loaded as a positive control. Following transfer, membranes were probed with the appropriate primary and secondary antibodies.
ACKNOWLEDGEMENTS
We thank E. Jacobs and L. Gao for technical assistance in generating the initial SPN cDNA clones. Special thanks go to the members of the Mieyal lab, particularly D. Starke for help with the gel filtration chromatography. We are also indebted to U. Fischer, K. Weis, R. Davis, H. Salz, J. Steitz and G. Dreyfuss for generously providing us with reagents. We are thankful to J. Valcarcel and P. Foersch at EMBL for helpful advice. We acknowledge the National Cell Culture Center for providing HeLaS9 cells. This work was supported by NIH grants R01-GM53034 and R01-NS41617 (to A.G.M.) and by a research grant from the Muscular Dystrophy Association. J.K.O. was supported in part by an NIH predoctoral traineeship (T32-GM08613). MDH was supported in part by an NIH post-doctoral fellowship (T32-HD07518) and by a career Development Grant from the Muscular Dystrophy Association.
REFERENCES
- 1.Görlich D, Mattaj IW. Nucleocytoplasmic transport. Science. 1996;271:1513–1518. doi: 10.1126/science.271.5255.1513. [DOI] [PubMed] [Google Scholar]
- 2.Allen TD, Cronshaw JM, Bagley S, Kiseleva E, Goldberg MW. The nuclear pore complex: mediator of translocation between nucleus and cytoplasm. J. Cell Sci. 2000;113:1651–1659. doi: 10.1242/jcs.113.10.1651. [DOI] [PubMed] [Google Scholar]
- 3.Ribbeck K, Görlich D. Kinetic analysis of translocation through nuclear pore complexes. EMBO J. 2001;20:1320–1330. doi: 10.1093/emboj/20.6.1320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Smith AE, Slepchenko BM, Schaff JC, Loew LM, Macara IG. Systems analysis of Ran transport. Science. 2002;295:488–491. doi: 10.1126/science.1064732. [DOI] [PubMed] [Google Scholar]
- 5.Ryan KJ, Wente SR. The nuclear pore complex: a protein machine bridging the nucleus and cytoplasm. Curr. Opin. Cell Biol. 2000;12:361–371. doi: 10.1016/s0955-0674(00)00101-0. [DOI] [PubMed] [Google Scholar]
- 6.Rout MP, Aitchison JD. The nuclear pore complex as a transport machine. J. Biol. Chem. 2001;276:16593–16596. doi: 10.1074/jbc.R100015200. [DOI] [PubMed] [Google Scholar]
- 7.Ström AC, Weis K. Importin-beta-like nuclear transport receptors. Genome Biol. 2001;2:3008–1. doi: 10.1186/gb-2001-2-6-reviews3008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Izaurralde E, Kutay U, von Kobbe C, Mattaj IW, Gorlich D. The asymmetric distribution of the constituents of the Ran system is essential for transport into and out of the nucleus. EMBO J. 1997;16:6535–6547. doi: 10.1093/emboj/16.21.6535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kalab P, Weis K, Heald R. Visualization of a Ran-GTP gradient in interphase and mitotic Xenopus egg extracts. Science. 2002;295:2452–2456. doi: 10.1126/science.1068798. [DOI] [PubMed] [Google Scholar]
- 10.Dingwall C, Laskey R. Nuclear targeting sequences—a consensus? Trends Biochem. Sci. 1991;16:478–481. doi: 10.1016/0968-0004(91)90184-w. [DOI] [PubMed] [Google Scholar]
- 11.Palacios I, Hetzer M, Adam SA, Mattaj IW. Nuclear import of U snRNPs requires importin beta. EMBO J. 1997;16:6783–6792. doi: 10.1093/emboj/16.22.6783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pasquinelli AE, Ernst RK, Lund E, Grimm C, Zapp ML, Rekosh D, Hammarskjold ML, Dahlberg JE. The constitutive transport element (CTE) of Mason-Pfizer monkey virus (MPMV) accesses a cellular mRNA export pathway. EMBO J. 1997;16:7500–7510. doi: 10.1093/emboj/16.24.7500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jullien D, Gorlich D, Laemmli UK, Adachi Y. Nuclear import of RPA in Xenopus egg extracts requires a novel protein XRIPalpha but not importin alpha. EMBO J. 1999;18:4348–4358. doi: 10.1093/emboj/18.15.4348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Conti E, Izaurralde E. Nucleocytoplasmic transport enters the atomic age. Curr. Opin. Cell Biol. 2001;13:310–319. doi: 10.1016/s0955-0674(00)00213-1. [DOI] [PubMed] [Google Scholar]
- 15.Will CL, Lührmann R. Spliceosomal U snRNP biogenesis, structure and function. Curr. Opin. Cell Biol. 2001;13:290–301. doi: 10.1016/s0955-0674(00)00211-8. [DOI] [PubMed] [Google Scholar]
- 16.Ohno M, Segref A, Bachi A, Wilm M, Mattaj IW. PHAX, a mediator of U snRNA nuclear export whose activity is regulated by phosphorylation. Cell. 2000;101:187–198. doi: 10.1016/S0092-8674(00)80829-6. [DOI] [PubMed] [Google Scholar]
- 17.Paushkin S, Gubitz AK, Massenet S, Dreyfuss G. The SMN complex, an assemblyosome of ribonucleoproteins. Curr. Opin. Cell Biol. 2002;14:305–312. doi: 10.1016/s0955-0674(02)00332-0. [DOI] [PubMed] [Google Scholar]
- 18.Mattaj IW. Cap trimethylation of U snRNA is cytoplasmic and dependent on U snRNP protein binding. Cell. 1986;46:905–911. doi: 10.1016/0092-8674(86)90072-3. [DOI] [PubMed] [Google Scholar]
- 19.Mouaikel J, Verheggen C, Bertrand E, Tazi J, Bordonné R. Hypermethylation of the cap structure of both yeast snRNAs and snoRNAs requires a conserved methyltransferase that is localized to the nucleolus. Mol. Cell. 2002;9:891–901. doi: 10.1016/s1097-2765(02)00484-7. [DOI] [PubMed] [Google Scholar]
- 20.Verheggen C, Lafontaine DLJ, Samarsky D, Mouaikel J, Blanchard J-M, Bordonné R, Bertrand E. Mammalian and yeast U3 snoRNPs are matured in specific and related nuclear compartments. EMBO J. 2002;21:2736–2745. doi: 10.1093/emboj/21.11.2736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mattaj IW, De Robertis EM. Nuclear segregation of U2 snRNA requires binding of specific snRNP proteins. Cell. 1985;40:111–118. doi: 10.1016/0092-8674(85)90314-9. [DOI] [PubMed] [Google Scholar]
- 22.Hamm J, Darzynkiewicz E, Tahara SM, Mattaj IW. The trimethylguanosine cap structure of U1 snRNA is a component of a bipartite nuclear targeting signal. Cell. 1990;62:569–577. doi: 10.1016/0092-8674(90)90021-6. [DOI] [PubMed] [Google Scholar]
- 23.Fischer U, Sumpter V, Sekine M, Satoh T, Lührmann R. Nucleo-cytoplasmic transport of U snRNPs: definition of a nuclear location signal in the Sm core domain that binds a transport receptor independently of the m3G cap. EMBO J. 1993;12:573–583. doi: 10.1002/j.1460-2075.1993.tb05689.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Huber J, Cronshagen U, Kadokura M, Marshallsay C, Wada T, Sekine M, Lührmann R. Snurportin1, an m3G-cap-specific nuclear import receptor with a novel domain structure. EMBO J. 1998;17:4114–4126. doi: 10.1093/emboj/17.14.4114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Paraskeva E, Izaurralde E, Bischoff FR, Huber J, Kutay U, Hartmann E, Lührmann R, Gorlich D. CRM1-mediated recycling of snurportin 1 to the cytoplasm. J. Cell Biol. 1999;145:255–264. doi: 10.1083/jcb.145.2.255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Huber J, Dickmanns A, Lührmann R. The importin-beta binding domain of snurportin1 is responsible for the Ran- and energy-independent nuclear import of spliceosomal U snRNPs in vitro. J. Cell Biol. 2002;156:467–479. doi: 10.1083/jcb.200108114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Fischer U, Lührmann R. An essential signaling role for the m3G cap in the transport of U1 snRNP to the nucleus. Science. 1990;249:786–790. doi: 10.1126/science.2143847. [DOI] [PubMed] [Google Scholar]
- 28.Fischer U, Heinrich J, van Zee K, Fanning E, Lührmann R. Nuclear transport of U1 snRNP in somatic cells: differences in signal requirement compared with Xenopus laevis oocytes. J. Cell Biol. 1994;125:971–980. doi: 10.1083/jcb.125.5.971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Matera AG. Nuclear bodies: multifaceted subdomains of the interchromatin space. Trends Cell Biol. 1999;9:302–309. doi: 10.1016/s0962-8924(99)01606-2. [DOI] [PubMed] [Google Scholar]
- 30.Gall JG. Cajal bodies: the first 100 years. Annu. Rev. Cell Dev. Biol. 2000;16:273–300. doi: 10.1146/annurev.cellbio.16.1.273. [DOI] [PubMed] [Google Scholar]
- 31.Sleeman JE, Lamond AI. Newly assembled snRNPs associate with coiled bodies before speckles, suggesting a nuclear snRNP maturation pathway. Curr. Biol. 1999;9:1065–1074. doi: 10.1016/s0960-9822(99)80475-8. [DOI] [PubMed] [Google Scholar]
- 32.Frey MR, Matera AG. Coiled bodies contain U7 small nuclear RNA and associate with specific DNA sequences in interphase cells. Proc. Natl Acad. Sci. USA. 1995;92:5915–5919. doi: 10.1073/pnas.92.13.5915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Smith K, Carter K, Johnson C, Lawrence J. U2 and U1 snRNA gene loci associate with coiled bodies. J. Cell. Biochem. 1995;59:473–485. doi: 10.1002/jcb.240590408. [DOI] [PubMed] [Google Scholar]
- 34.Jacobs EY, Frey MR, Wu W, Ingledue TC, Gebuhr TC, Gao L, Marzluff WF, Matera AG. Coiled bodies preferentially associate with U4, U11, and U12 small nuclear RNA genes in interphase HeLa cells but not with U6 and U7 genes. Mol. Biol. Cell. 1999;10:1653–1663. doi: 10.1091/mbc.10.5.1653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Frey MR, Bailey AD, Weiner AM, Matera AG. Association of snRNA genes with coiled bodies is mediated by nascent snRNA transcripts. Curr. Biol. 1999;9:126–131. doi: 10.1016/s0960-9822(99)80066-9. [DOI] [PubMed] [Google Scholar]
- 36.Frey MR, Matera AG. RNA-mediated interaction of Cajal bodies and U2 snRNA genes. J. Cell Biol. 2001;154:499–509. doi: 10.1083/jcb.200105084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Marshallsay C, Lührmann R. In vitro nuclear import of snRNPs: cytosolic factors mediate m3G-cap dependence of U1 and U2 snRNP transport. EMBO J. 1994;13:222–231. doi: 10.1002/j.1460-2075.1994.tb06252.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Fischer U, Liu Q, Dreyfuss G. The SMN-SIP1 complex has an essential role in spliceosomal snRNP biogenesis. Cell. 1997;90:1023–1029. doi: 10.1016/s0092-8674(00)80368-2. [DOI] [PubMed] [Google Scholar]
- 39.Gangwani L, Mikrut M, Theroux S, Sharma M, Davis RJ. Spinal muscular atrophy disrupts the interaction of ZPR1 with the survival motor neurons protein. Nat. Cell Biol. 2001;3:376–383. doi: 10.1038/35070059. [DOI] [PubMed] [Google Scholar]
- 40.Feldherr CM, Akin D. EM visualization of nucleocytoplasmic transport processes. Electron Microsc. Rev. 1990;3:73–86. doi: 10.1016/0892-0354(90)90014-j. [DOI] [PubMed] [Google Scholar]
- 41.Eriksson SA, Mannervik B. The reduction of the L-cysteine-glutathione mixed disulfide in rat liver. Involvement of an enzyme catalyzing thiol-disulfide interchange. FEBS Lett. 1970;7:26–28. doi: 10.1016/0014-5793(70)80608-1. [DOI] [PubMed] [Google Scholar]
- 42.Chan EKL, Takano S, Andrade LEC, Hamel JC, Matera AG. Structure, expression and chromosomal localization of the human p80-coilin gene. Nucleic Acids Res. 1994;22:4462–4469. doi: 10.1093/nar/22.21.4462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Charroux B, Pellizzoni L, Perkinson RA, Shevchenko A, Mann M, Dreyfuss G. Gemin3. A novel dead box protein that interacts with SMN, the spinal muscular atrophy gene product, and is a component of gems. J. Cell Biol. 1999;147:1181–1194. doi: 10.1083/jcb.147.6.1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gangwani L, Mikrut M, Galcheva-Gargova Z, Davis RJ. Interaction of ZPR1 with translation elongation factor-1 alpha in proliferating cells. J. Cell Biol. 1998;143:1471–1484. doi: 10.1083/jcb.143.6.1471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Galcheva-Gargova Z, Konstantinov KN, Wu IH, Klier FG, Barrett T, Davis RJ. Binding of zinc finger protein ZPR1 to the epidermal growth factor receptor. Science. 1996;272:1797–1802. doi: 10.1126/science.272.5269.1797. [DOI] [PubMed] [Google Scholar]
- 46.Galcheva-Gargova Z, Gangwani L, Konstantinov KN, Mikrut M, Theroux SJ, Enoch T, Davis RJ. The cytoplasmic zinc finger protein ZPR1 accumulates in the nucleolus of proliferating cells. Mol. Biol. Cell. 1998;9:2963–2971. doi: 10.1091/mbc.9.10.2963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hebert MD, Szymczyk PW, Shpargel KB, Matera AG. Coilin forms the bridge between Cajal bodies and SMN, the spinal muscular atrophy protein. Genes Dev. 2001;15:2720–2729. doi: 10.1101/gad.908401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sleeman JE, Ajuh P, Lamond AI. snRNP protein expression enhances the formation of Cajal bodies containing p80-coilin and SMN. J. Cell Sci. 2001;114:4407–4419. doi: 10.1242/jcs.114.24.4407. [DOI] [PubMed] [Google Scholar]
- 49.Meister G, Buhler D, Pillai R, Lottspeich F, Fischer U. A multiprotein complex mediates the ATP-dependent assembly of spliceosomal U snRNPs. Nat. Cell Biol. 2001;3:945–949. doi: 10.1038/ncb1101-945. [DOI] [PubMed] [Google Scholar]
- 50.Yong J, Pellizzoni L, Dreyfuss G. Sequence-specific interaction of U1 snRNA with the SMN complex. EMBO J. 2002;21:1188–1196. doi: 10.1093/emboj/21.5.1188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Pellizzoni L, Charroux B, Dreyfuss G. SMN mutants of spinal muscular atrophy patients are defective in binding to snRNP proteins. Proc. Natl Acad. Sci. USA. 1999;96:11167–11172. doi: 10.1073/pnas.96.20.11167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Bühler D, Raker V, Lührmann R, Fischer U. Essential role for the Tudor domain of SMN in spliceosomal U snRNP assembly: implications for spinal muscular atrophy. Hum. Mol. Genet. 1999;8:2351–2357. doi: 10.1093/hmg/8.13.2351. [DOI] [PubMed] [Google Scholar]
- 53.Charroux B, Pellizzoni L, Perkinson RA, Yong J, Shevchenko A, Mann M, Dreyfuss G. Gemin4: a novel component of the SMN complex that is found in both gems and nucleoli. J. Cell Biol. 2000;148:1177–1186. doi: 10.1083/jcb.148.6.1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Brahms H, Raymackers J, Union A, de Keyser F, Meheus L, Lührmann R. The C-terminal RG dipeptide repeats of the spliceosomal Sm proteins D1 and D3 contain symmetrical dimethylarginines, which form a major B-cell epitope for anti-Sm autoantibodies. J. Biol. Chem. 2000;275:17122–17129. doi: 10.1074/jbc.M000300200. [DOI] [PubMed] [Google Scholar]
- 55.Brahms H, Meheus L, de Brabandere V, Fischer U, Lührmann R. Symmetrical dimethylation of arginine residues in spliceosomal Sm protein B/B′and the Sm-like protein LSm4, and their interaction with the SMN protein. RNA. 2001;7:1531–1542. doi: 10.1017/s135583820101442x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Friesen WJ, Paushkin S, Wyce A, Massenet S, Pesiridis GS, Van Duyne G, Rappsilber J, Mann M, Dreyfuss G. The methylosome, a 20S complex containing JBP1 and pICln, produces dimethylarginine-modified Sm proteins. Mol. Cell. Biol. 2001;21:8289–8300. doi: 10.1128/MCB.21.24.8289-8300.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Friesen WJ, Massenet S, Paushkin S, Wyce A, Dreyfuss G. SMN, the product of the spinal muscular atrophy gene, binds preferentially to dimethylarginine-containing protein targets. Mol. Cell. 2001;7:1111–1117. doi: 10.1016/s1097-2765(01)00244-1. [DOI] [PubMed] [Google Scholar]
- 58.Meister G, Eggert C, Buhler D, Brahms H, Kambach C, Fischer U. Methylation of Sm proteins by a complex containing PRMT5 and the putative U snRNP assembly factor pICln. Curr. Biol. 2001;11:1990–1994. doi: 10.1016/s0960-9822(01)00592-9. [DOI] [PubMed] [Google Scholar]
- 59.Pellizzoni L, Kataoka N, Charroux B, Dreyfuss G. A novel function for SMN, the spinal muscular atrophy disease gene product, in pre-mRNA splicing. Cell. 1998;95:615–624. doi: 10.1016/s0092-8674(00)81632-3. [DOI] [PubMed] [Google Scholar]
- 60.Jones KW, Gorzynski K, Hales CM, Fischer U, Badbanchi F, Terns RM, Terns MP. Direct interaction of the spinal muscular atrophy disease protein SMN with the small nucleolar RNA-associated protein fibrillarin. J. Biol. Chem. 2001;276:38645–38651. doi: 10.1074/jbc.M106161200. [DOI] [PubMed] [Google Scholar]
- 61.Pellizzoni L, Baccon J, Charroux B, Dreyfuss G. The survival of motor neurons (SMN) protein interacts with the snoRNP proteins fibrillarin and GAR1. Curr. Biol. 2001;11:1079–1088. doi: 10.1016/s0960-9822(01)00316-5. [DOI] [PubMed] [Google Scholar]
- 62.Terns MP, Terns RM. Macromolecular complexes: SMN—the master assembler. Curr. Biol. 2001;11:R862–R864. doi: 10.1016/s0960-9822(01)00517-6. [DOI] [PubMed] [Google Scholar]
- 63.Gubitz AK, Mourelatos Z, Abel L, Rappsilber J, Mann M, Dreyfuss G. Gemin5, a novel WD repeat protein component of the SMN complex that binds Sm proteins. J. Biol. Chem. 2002;277:5631–5636. doi: 10.1074/jbc.M109448200. [DOI] [PubMed] [Google Scholar]
- 64.Lerner EA, Lerner MR, Janeway CA, Steitz JA. Monoclonal antibodies to nucleic acid-containing cellular constituents: probes for molecular biology and autoimmune disease. Proc. Natl Acad. Sci. USA. 1981;78:2737–2741. doi: 10.1073/pnas.78.5.2737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Andrade LEC, Chan EKL, Raska I, Peebles CL, Roos G, Tan EM. Human autoantibody to a novel protein of the nuclear coiled body: immunological characterization and cDNA cloning of p80 coilin. J. Exp. Med. 1991;173:1407–1419. doi: 10.1084/jem.173.6.1407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Dignam JD, Lebovitz RM, Roeder RG. Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res. 1983;11:1475–1489. doi: 10.1093/nar/11.5.1475. [DOI] [PMC free article] [PubMed] [Google Scholar]