

Abstract
Human DNA polymerase (Pol)κ is a member of the Y family of DNA polymerases. Unlike Polη, another member of this family, which carries out efficient translesion synthesis through various DNA lesions, the role of Polκ in lesion bypass has remained unclear. Recent studies, however, have indicated that Polκ is a proficient extender of mispaired primer termini on undamaged DNAs and also on cis-syn thymine-thymine (T-T) dimer-containing DNA. Here we determine whether Polκ can promote the efficient bypass of DNA lesions by extending from the nucleotides inserted opposite the lesion site by another DNA polymerase. From steady-state kinetic analyses, we find that Polκ is highly inefficient at incorporating nucleotides opposite an O6-methyl guanine (m6G) lesion, but it efficiently extends from the T or C nucleotide incorporated opposite this lesion by Polδ. Opposite an 8-oxoguanine (8-oxoG) lesion, Polκ efficiently inserts an A and then proficiently extends from it. Importantly, for both these DNA lesions, however, the most efficient bypass occurs when Polδ is combined with Polκ; in this reaction, Polκ performs the extension step after the incorporation of nucleotides opposite these lesion sites by Polδ. These studies reveal a role for Polκ in the extension phase of lesion bypass.
Although a variety of repair mechanisms exists to remove DNA lesions formed by the action of extrinsic and intrinsic DNA-damaging agents, lesions that escape repair can present a block to normal replication. Replication through such lesions, however, can be achieved by the action of translesion-synthesis DNA polymerases. In both yeast and humans, DNA polymerase (Pol)η promotes replication through UV-induced cyclobutane pyrimidine dimers (1, 2). Steady-state kinetic studies have indicated that Polη replicates through a cis-syn thymine-thymine (T-T) dimer with the same efficiency and accuracy as it replicates through undamaged Ts (3, 4), and genetic studies in yeast have also implicated Polη in the error-free bypass of cyclobutane pyrimidine dimers formed at TC and CC sites (5). Because of the involvement of Polη in the error-free bypass of cyclobutane pyrimidine dimers, its mutational inactivation in humans (2, 6) causes an increase in the incidence of UV mutagenesis (7, 8) and leads to the cancer-prone syndrome, the variant form of xeroderma pigmentosum (XP-V). Polη can replicate efficiently through other DNA lesions as well such as 7,8-dihydro-8-oxoguanine (8-oxoG) and O6-methyl guanine (m6G) (9, 10).
By contrast to Polη, the ability of which to replicate through DNA lesions derives both from its proficient ability to insert nucleotides opposite the lesion site and to extend proficiently from the inserted nucleotide, the bypass of certain DNA lesions requires the sequential action of two DNA polymerases, in which one inserts the nucleotide opposite the lesion and the other extends from the inserted nucleotide. Translesion synthesis through DNA lesions such as a (6-4) T-T photoproduct or an abasic (AP) site requires the consecutive action of two DNA polymerases, wherein Polζ acts at the extension step (11). Steady-state kinetic studies with yeast Polζ have indicated that it is very efficient at extending from mispaired primer termini on undamaged DNAs (11). Polζ is also proficient in extending from nucleotides opposite DNA lesions; for example, Polζ proficiently extends from a G opposite the 3′T of a (6-4) T-T photoproduct (11, 12) and from an A opposite an AP site (13).
In addition to Polη and Polζ, humans contain two other DNA polymerases, Polι and Polκ. Whereas Polη, Polι, and Polκ are members of the Y family of DNA polymerases, Polζ is unrelated to these polymerases and is a member of the Polα family. Polι shows little proficiency for replicating through a cis-syn T-T dimer, but it is able to incorporate nucleotides opposite the 3′T of the (6-4) T-T photoproduct and also opposite an AP site (11), suggesting a role for this polymerase at the insertion step of lesion bypass. Polκ (originally called Polθ by us), however, shows no tendency to replicate through a cis-syn T-T dimer or a (6-4) T-T photoproduct, and it is quite inefficient at replicating through an AP site (14). Polκ is also highly inefficient at replicating past an acetylaminofluorene (15), a 1,N6-ethenodeoxyadenosine (16), or a benzo[a]pyrene-derived DNA adduct (17). Although Polκ is unable to efficiently replicate through the many DNA lesions that have been examined thus far, our recent observations indicating that Polκ is a promiscuous extender of mispaired primer termini on undamaged DNAs (18) have suggested that it could function in the extension of such primer termini formed during normal DNA replication. Polκ also can extend proficiently from a G nucleotide opposite the 3′T of a T-T dimer, but it is highly inefficient in inserting nucleotides opposite the 3′T of this lesion (18).
Here, we examine whether Polκ can promote the efficient bypass of DNA lesions by extending from the nucleotides inserted opposite the lesion site by another DNA polymerase. For the two DNA lesions examined, m6G and 8-oxoG, from steady-state kinetic analyses we determine that Polκ is very inefficient at inserting nucleotides opposite m6G, but it proficiently extends from a T or a C nucleotide placed opposite this lesion. Opposite an 8-oxoG lesion, Polκ efficiently inserts an A and then proficiently extends from it. For both these DNA lesions, however, the most efficient bypass occurs when the replicative polymerase Polδ is combined with Polκ. In this reaction, Polκ promotes efficient lesion bypass by extending from the nucleotides incorporated opposite these lesion sites by Polδ.
Materials and Methods
Proteins and DNA Substrates.
Human Polκ was expressed in and purified from yeast strain BJ5464 as described (14). Human Polδ, which was a gift from J. Hurwitz (Memorial Sloan–Kettering Cancer Center, New York), was purified as described (19). Oligonucleotides were synthesized by Midland Certified Reagents (Midland, TX). DNA substrates S1- (shown in Fig. 1) were generated by annealing a 75-nt oligomer template, 5′-AGC TAC CAT GCC TGC CTC AAG AAT TCG TAA XAT GCC TAC ACT GGA GTA CCG GAG CAT CGT CGT GAC TGG GAA AAC-3′, containing a G (S1-G), an m6G (S1-m6G), or a 7,8-dihydro-8-oxoG (S1–8-oxoG) at position 31 (X) to the 5′ 32P-labeled oligonucleotide primer N4264 (5′-GTT TTC CCA GTC ACG ACG ATG CTC CGG TAC TCC AGT GTA G-3′). For DNA substrate S2- (shown in Fig. 2A and also used for steady-state kinetic analysis of insertion reactions), the 75-nt templates used for generating the substrates S1- were annealed to the 5′ 32P-labeled oligonucleotide primer N4309 (5′-GTT TTC CCA GTC ACG ACG ATG CTC CGG TAC TCC AGT GTA GGC AT-3′), yielding substrates S2-G, S2–8-oxoG, and S2-m6G. DNA substrates S3- (shown in Fig. 2B and used for steady-state kinetic analysis of extension reactions in Fig. 3B) were generated by annealing the 75-nt oligomer templates used for generating the substrates S1- to the 5′ 32P-labeled oligonucleotide primers, 5′-GTT TTC CCA GTC ACG ACG ATG CTC CGG TAC TCC AGT GTA GGCATN-3′, where N is G, A, T, or C.
Fig 1.

DNA synthesis across an m6G or 8-oxoG lesion by human Polδ and Polκ. Sequences adjacent to the primer–template junction are shown for the 5′ 32P-labeled primer and the 75-nt template. The position corresponding to the undamaged G, m6G, or 8-oxoG on the template is indicated by G*. The DNA substrate (20 nM) was incubated with human Polκ (3 nM), human Polδ (0.03 unit), or a mixture of these two enzymes in the presence of 5 or 100 μM of each of four dNTPs in 10-μl reactions at 37°C for 10 min. The reaction products were resolved on a 10% denaturing polyacrylamide gel and visualized by autoradiography. The gel was analyzed by PhosphorImager, and the amounts of the products of synthesis past the undamaged G, m6G, or 8-oxoG are indicated.
Fig 2.

Nucleotide incorporation opposite the m6G and 8-oxoG lesions by human Polδ and extension from the ensuing primer ends by human Polκ. (A) Specificity of deoxynucleotide incorporation by human Polδ opposite the m6G and 8-oxoG lesions. Nucleotide sequences adjacent to the primer–template junction are shown. Human Polδ (0.03 unit) was incubated with the DNA substrate (20 nM) in the presence of 100 μM of a single (G, A, T, or C) deoxynucleotide for 10 min at 37°C. (B) Extension of primers with varying 3′ ends opposite the m6G and 8-oxoG lesions by human Polκ. Each of the four different primers, differing only in the 3′-terminal nucleotide, was annealed to the template oligonucleotide containing an undamaged G, an m6G, or an 8-oxoG residue at the same position. In the primer template shown, N designates the position of the variable terminal primer nucleotide and G* designates the position of a G, an m6G, or an 8-oxoG residue. DNA substrate (20 nM) was incubated with human Polκ (3 nM) in the presence of each of the four dNTPs (100 μM) at 37°C for 10 min. The gel was analyzed by PhosphorImager, and the amount of the product of primer extension opposite from the undamaged G, m6G, or 8-oxoG is indicated.
Fig 3.

Insertion and extension reactions catalyzed by human Polκ on 8-oxoG- and m6G-containing DNA templates. (A) Deoxynucleotide incorporation across from a template 8-oxoG or m6G residue. A portion of the DNA substrate is shown, and the position of an m6G, 8-oxoG, or undamaged G residue in the template oligonucleotide is indicated by an asterisk. Polκ (1 nM) was incubated with the primer-template DNA substrate (10 nM) and increasing concentrations (0–1,000 μM) of a single deoxynucleotide (dGTP, dATP, dTTP, or dCTP) in standard reaction buffer for 10 min at 37°C. As a control reaction, the incorporation of dCTP (0–40 μM) opposite an undamaged G residue in the template was carried out. The quenched samples were analyzed by 10% denaturing PAGE. (B) Extension of primers containing a G, A, T, or C residue opposite an m6G or 8-oxoG residue in the template by Polκ. Primers differing only in the last nucleotide at the 3′ end were annealed separately to the m6G- or 8-oxoG-containing template as shown. As a control, the extension of primer containing a C opposite an undamaged G residue in the template was examined. Reactions were performed in the presence of increasing dTTP concentrations (0–1,000 or 0–40 μM) as described for A.
DNA-Polymerase Assays.
A standard DNA-polymerase reaction (10 μl) contained 40 mM Tris⋅HCl (pH 7.5), 8 mM MgCl2, 1 mM DTT, 100 μg/ml BSA, 10% glycerol, 20 nM 5′ 32P-labeled oligonucleotide primer annealed to an oligonucleotide template, and 5 or 100 μM of each of all four deoxynucleotides (dGTP, dATP, dTTP, and dCTP) as indicated in the figure legends. Reactions were started by the addition of Polκ (3 nM) or Polδ (0.03 unit, unit is as defined in ref. 19) and incubated at 37°C for 10 min followed by quenching by the addition of loading buffer (40 μl) containing 20 mM EDTA, 95% formamide, 0.3% bromophenol blue, and 0.3% cyanol blue. The reaction products were resolved on 10% polyacrylamide gels containing 8 M urea. Quantitation of the results was done by using a Molecular Dynamics STORM PhosphorImager and IMAGEQUANT software. The concentration of the product of the bypass reaction was calculated as the ratio of the intensity of bands that lie above the lesion site to the intensity of all bands including the primer.
Analysis of Steady-State Kinetics.
Steady-state kinetic analyses for deoxynucleotide incorporation opposite an 8-oxoG or m6G or primer extension opposite from these lesions were performed as described (20–22). Briefly, Polκ (1 nM) was incubated with 10 nM DNA substrate in the presence of increasing concentrations of a single deoxynucleotide for 10 min. The reaction products were resolved on 15% polyacrylamide gels containing 8 M urea. Gel-band intensities of the substrates and products were quantitated by PhosphorImager, and the observed rate of deoxynucleotide incorporation was plotted as a function of dNTP concentration. The data were fit by nonlinear regression using SIGMAPLOT 5.0 to the Michaelis–Menten equation describing a hyperbola, v = (Vmax × [dNTP]/(Km + [dNTP]). The kcat and Km steady-state parameters were obtained from the fit and used to calculate the frequency of deoxynucleotide incorporation (finc) and the intrinsic efficiency of mispair extension (f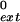 ) for each primer/template pair by using the equation finc or f
) for each primer/template pair by using the equation finc or f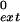 = (kcat/Km)mispaired/(kcat/Km)paired.
= (kcat/Km)mispaired/(kcat/Km)paired.
Results
Replication Through the m6G and 8-oxoG Lesions by the Concerted Action of Human Polδ and Polκ.
We examined the ability of human Polκ to replicate past an m6G and an 8-oxoG lesion in the template DNA, and we also carried out bypass reactions by human Polδ and by the combination of Polδ and Polκ. Running-start experiments were performed by using DNA substrates constructed by annealing a linear 75-nt template DNA containing a single m6G, an 8-oxoG, or, as a control, a G residue 45 nt from the 3′ end to a 5′ 32P-labeled 40-nt primer (Fig. 1), and DNA-synthesis reactions were carried out at low (5 μM) and high (100 μM) nucleotide concentrations. The m6G lesion is a strong block to DNA synthesis, and even at 100 μM dNTP, Polκ replicated through only ≈14% (Fig. 1, lane 10), and Polδ replicated through only ≈4% of this lesion (Fig. 1, lane 12) compared with synthesis on template containing an undamaged G residue (Fig. 1, lanes 3 and 5). Furthermore, Polκ exhibited a strong stall site right before the m6G residue, indicating a strong inhibition of insertion across from this lesion by this polymerase (Fig. 1, lanes 9 and 10). Although Polδ showed some ability to insert nucleotides opposite the m6G lesion, it was not able to subsequently extend from the resulting primer end (Fig. 1, lane 12). Remarkably, however, the combination of Polκ with Polδ promoted m6G bypass, and together they replicated through the m6G lesion ≈50% as efficiently as through the undamaged G residue (Fig. 1, compare lanes 13 and 14 with lanes 6 and 7, respectively).
Polκ replicated through an 8-oxoG residue with ≈50% the efficiency of replicating through an undamaged G (Fig. 1, compare lanes 16 and 17 with lanes 2 and 3, respectively). Moreover, Polκ did not exhibit a prominent stall site just before the 8-oxoG residue or opposite this lesion, suggesting it is able to insert nucleotides opposite the 8-oxoG lesion efficiently and to extend from the inserted nucleotide. Polδ can also replicate through an 8-oxoG lesion, but it does so 3- to 4-fold less efficiently than it replicates through an undamaged G (Fig. 1, compare lanes 18 and 19 with lanes 4 and 5, respectively.) Importantly, however, the most striking increase in 8-oxoG bypass occurred when Polδ was combined with Polκ, and at 5 μM dNTP, the two polymerases together replicated through the 8-oxoG lesion as effectively as through the undamaged G (Fig. 1, compare lanes 20 and 6). The key observation here is that Polδ and Polκ together are much more effective in m6G and 8-oxoG bypass than either polymerase alone. These observations suggested that because of its proximity to the lesion site, Polδ would contribute to the insertion of nucleotides opposite these lesions, whereas Polκ could carry out the subsequent extension from the ensuing primer end.
Nucleotide Incorporation Opposite m6G and 8-oxoG by Human Polδ.
We have shown previously that yeast Polδ inserts a T or C opposite an m6G (10) and an A opposite an 8-oxoG lesion (9), promoting the mutagenic bypass of these lesions. To examine whether human Polδ incorporates the same nucleotides opposite these lesions as yeast Polδ, DNA substrates containing an undamaged G, an m6G, or an 8-oxoG residue next to the primer–template junction were incubated with human Polδ in the presence of just one dNTP. Human Polδ inserted a T or C opposite the m6G lesion (Fig. 2A, lanes 7 and 8), whereas opposite the 8-oxoG lesion, it predominantly inserted an A (Fig. 2A, lane 10), but some C nucleotide was inserted also (Fig. 2A, lane 12).
Extension of Primer Ends Opposite from the m6G and 8-oxoG Lesions by Human Polκ.
Next we examined the ability of Polκ to extend from the various 3′-primer termini situated across from the m6G or 8-oxoG lesion in the 75-nt template in the presence of 100 μM of each of the four dNTPs (Fig. 2B). Polκ extended from the T and C 3′-primer ends situated opposite the m6G site with ≈70% of the efficiency of extension from the C 3′-primer end situated opposite undamaged G (Fig. 2B, compare lanes 7 and 8 with lane 4). The extension from the A or C 3′-primer end opposite from the 8-oxoG residue was efficient also, occurring with ≈70% the efficiency of extension from the C 3′-primer end situated opposite undamaged G (Fig. 2B, compare lanes 10 and 12 with lane 4). Thus, after the insertion of nucleotides opposite the m6G or 8-oxoG lesion by Polδ, Polκ could subsequently extend from the inserted nucleotides.
Steady-State Kinetic Analyses of Nucleotide Insertion and Extension Reactions Across from the m6G and 8-oxoG Lesions by Human Polκ.
To characterize further the role of Polκ in the bypass of m6G and 8-oxoG lesions, we measured the steady-state kinetic parameters of nucleotide insertion and extension reactions opposite from these lesions. The kinetics of insertion of a single deoxynucleotide opposite an m6G and an 8-oxoG (Fig. 3A) and the kinetics of the addition of the next correct nucleotide to various 3′-primer termini situated across from m6G and 8-oxoG (Fig. 3B) were determined as a function of deoxynucleotide concentration under steady-state conditions. We incubated Polκ with m6G- or 8-oxoG-containing DNA substrates and various concentrations of a single nucleotide. The rate of nucleotide incorporation was plotted as a function of nucleotide concentration. The steady-state apparent kcat and Km values for each nucleotide incorporation and for extension of each primer terminus were obtained from the curve fitted to the Michaelis–Menten equation by using nonlinear regression. The frequency of nucleotide incorporation, finc, and the relative efficiency of extension, f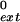 , were calculated as the ratio of the efficiency (kcat/Km) of incorrect nucleotide incorporated or extended from to the efficiency (kcat/Km) of correct nucleotide incorporated or extended from, respectively (Tables 1 and 2).
, were calculated as the ratio of the efficiency (kcat/Km) of incorrect nucleotide incorporated or extended from to the efficiency (kcat/Km) of correct nucleotide incorporated or extended from, respectively (Tables 1 and 2).
Table 1.
Steady-state kinetic parameters of nucleotide-insertion reactions opposite m6G and 8-oxoG template residues by human Polκ
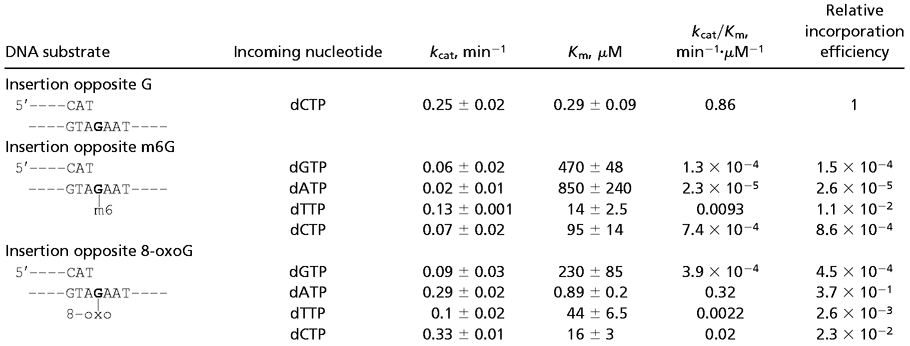 |
Table 2.
Steady-state kinetic parameters of extension reactions opposite from m6G and 8-oxoG catalyzed by human Polκ
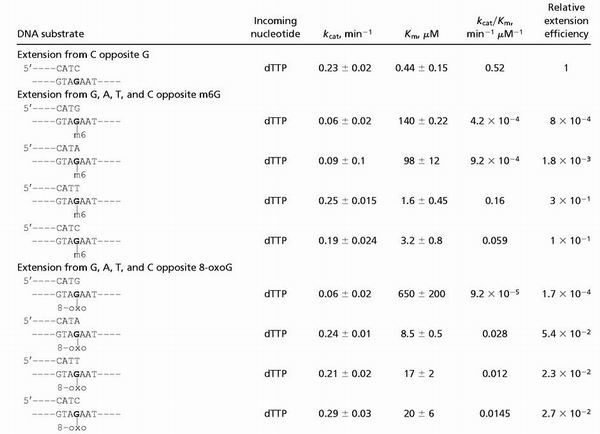 |
As indicated by the kcat/Km values shown in Table 1, Polκ incorporated nucleotides opposite the m6G lesion inefficiently. Compared with the incorporation of a C opposite the undamaged G-template residue, Polκ incorporated nucleotides opposite m6G with frequencies ranging from 1 × 10−2 to 2.6 × 10−5. Polκ incorporated a T ≈13-fold more efficiently than C opposite m6G; however, even T incorporation opposite m6G was ≈100-fold less efficient than the C incorporation opposite undamaged G (Table 1). In contrast, Polκ extended opposite from the m6G lesion quite efficiently. As shown in Table 2, compared with the extension from a C opposite an undamaged G in the template, Polκ extended from a T and a C opposite from m6G ≈3- and 10-fold less efficiently, respectively. The extension from the A and G nucleotides opposite m6G was much less efficient.
Polκ is quite efficient at inserting an A opposite the 8-oxoG lesion (Table 1). Compared with the incorporation of a C opposite undamaged G in the template, Polκ incorporated an A opposite 8-oxoG only ≈3-fold less efficiently. Polκ also can insert a C opposite 8-oxoG, but it is ≈40-fold less efficient at it than the insertion of C opposite the undamaged G-template residue. In addition, Polκ is an efficient extender opposite from the 8-oxoG lesion (Table 2). Compared with the extension from a C opposite an undamaged G in the template, Polκ extended from an A, C, or T opposite 8-oxoG only ≈19-, 36-, and 43-fold less efficiently, respectively. Overall, Polκ is quite proficient in inserting an A opposite 8-oxoG as well as in extending from this nucleotide.
Discussion
Here we show that although neither Polδ or Polκ is able to replicate through an m6G lesion, the combination of these two polymerases performs this task quite effectively. Although Polδ can insert a T or C residue opposite an m6G lesion, it is very poor at extending from the inserted nucleotide. Polκ, on the other hand, is highly inefficient at inserting nucleotides opposite m6G, but it extends from the T and C nucleotides incorporated opposite this DNA lesion quite efficiently. Efficient replication through an m6G lesion then could be achieved by the sequential action of Polδ and Polκ, with Polδ acting at the insertion step and Polκ at the extension step.
Although both Polδ and Polκ are able to replicate through an 8-oxoG lesion by primarily inserting an A opposite it, for this lesion also the two polymerases together are much more effective in bypassing it. From steady-state kinetic analyses we find that Polκ is more efficient at extending from an A than from a C opposite from 8-oxoG. Thus, after the incorporation of an A, or less frequently of a C, opposite 8-oxoG by Polδ, Polκ could carry out the subsequent extension step. Because during DNA replication Polδ would be the first polymerase to encounter the DNA lesion, we expect it to make a significant contribution to the nucleotide-incorporation step opposite the m6G and 8-oxoG lesions, after which Polκ could contribute to the extension step. Thus, Polκ could promote efficient replication through m6G and 8-oxoG lesions by extending from the nucleotides inserted opposite these lesions by Polδ.
Human Polδ seems to incorporate nucleotides opposite m6G and 8-oxoG lesions rather weakly, and this is evidenced by the presence of a strong stall site just before these lesion sites. The apparently weak nucleotide-incorporation reaction by Polδ opposite these lesion sites may derive from its 3′ → 5′ exonuclease activity, which in the absence of proficient extension could remove the inserted nucleotide. In the presence of Polκ, however, because of its ability to extend efficiently from the primer terminus, the forward synthesis reaction would prevail over the backward exonucleolytic reaction of Polδ, enabling the proficient bypass of these DNA lesions by the combination of Polδ and Polκ.
Genetic and biochemical studies in yeast have indicated a role for both Polδ and Polη in promoting replication through an m6G lesion (10); however, Polη is much more efficient in replicating through this DNA lesion than Polδ. Human Polη is also able to replicate through the m6G lesion by inserting a T or C and by extending from the inserted nucleotide (10). In Fig. 4A we present various routes by which replication through the m6G lesion could be accomplished in human cells. Although we find that human Polδ is quite ineffective in replicating through this DNA lesion, the possibility that it contributes to some of the lesion bypass, particularly when it is together with the accessory proteins PCNA, RFC, and RPA during its role in replication, cannot be excluded. However, the more likely routes by which m6G bypass could occur in human cells are the following. (i) Subsequent to the insertion of a T or C by Polδ, the extension step is mediated by Polκ or by Polη. Because of the efficient ability of Polη and Polκ to extend from the T or C residue opposite from m6G, we expect both these polymerases to compete at this step. (ii) Alternatively, Polη carries out both the insertion and extension steps.
Fig 4.

Alternate routes of DNA synthesis through an m6G (A) or 8-oxoG (B) lesion in human cells. For both these DNA lesions, after the insertion of nucleotides by Polδ, the extension step could be performed by Polκ. Alternatively, Polη could carry out lesion bypass by executing both the insertion and extension steps, or subsequent to nucleotide insertion by Polδ, Polη conducts the extension step. Although Polδ could replicate through these DNA lesions, we expect this to make a relatively small contribution to lesion bypass. The more frequently inserted nucleotides are indicated in bold.
Although some replication through the 8-oxoG lesion in human cells could occur by the action of Polδ, we expect the other pathways to play a more predominant role in the bypass of this lesion also (Fig. 4B). Thus, replication through this lesion could entail the sequential action of Polδ and Polκ, in which Polδ inserts the nucleotide opposite 8-oxoG and Polκ subsequently extends; alternatively, Polη could replicate through the lesion by executing both the steps. Although yeast Polη predominantly inserts a C opposite an 8-oxoG and efficiently extends from it, human Polη bypasses this lesion less accurately, because in addition to a C, it also inserts some A (9). Polη could contribute additionally to the extension from nucleotides inserted opposite 8-oxoG by Polδ.
Polκ is much more limited in its ability to bypass DNA lesions than Polη. Thus, whereas Polη efficiently inserts the correct nucleotide opposite the 3′T of a T-T dimer and proficiently extends from the inserted nucleotide, Polκ is unable to insert nucleotides opposite the 3′T of this lesion; however, it can extend efficiently from a G or A nucleotide placed opposite the 3′T of a T-T dimer. Opposite 8-oxoG and m6G lesions also, Polη is able both to insert nucleotides and to extend, whereas Polκ can perform only the extension step for the m6G lesion and both the insertion and extension steps for the 8-oxoG lesion. From the x-ray crystal structure of Polη with modeled DNA and an incoming dNTP, it has been inferred that the Polη active site is quite open and can accommodate two template nucleotides: the templating residue and the next 5′-unpaired residue (23). The more limited ability of Polκ than that of Polη to insert nucleotides opposite DNA lesions may result from the more intimate contacts of Polκ with the templating nucleotide and the incoming dNTP. The proficient ability of Polκ to extend from mispaired primer termini on undamaged DNAs and from the primer termini opposite certain DNA lesions, however, may reflect that this polymerase is fairly tolerant of geometric distortions at the primer terminus.
Acknowledgments
This work was supported by National Institutes of Health Grants GM19261 and CA80882.
Abbreviations
Pol, DNA polymerase
8-oxoG, 8-oxoguanine
m6G, O6-methyl guanine
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Johnson R. E., Prakash, S. & Prakash, L. (1999) Science 283, 1001-1004. [DOI] [PubMed] [Google Scholar]
- 2.Masutani C., Kusumoto, R., Yamada, A., Dohmae, N., Yokoi, M., Yuasa, M., Araki, M., Iwai, S., Takio, K. & Hanaoka, F. (1999) Nature 399, 700-704. [DOI] [PubMed] [Google Scholar]
- 3.Johnson R. E., Washington, M. T., Prakash, S. & Prakash, L. (2000) J. Biol. Chem. 275, 7447-7450. [DOI] [PubMed] [Google Scholar]
- 4.Washington M. T., Johnson, R. E., Prakash, S. & Prakash, L. (2000) Proc. Natl. Acad. Sci. USA 97, 3094-3099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Yu S.-L., Johnson, R. E., Prakash, S. & Prakash, L. (2001) Mol. Cell. Biol. 21, 185-188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Johnson R. E., Kondratick, C. M., Prakash, S. & Prakash, L. (1999) Science 285, 263-265. [DOI] [PubMed] [Google Scholar]
- 7.Wang Y.-C., Maher, V. M., Mitchell, D. L. & McCormick, J. J. (1993) Mol. Cell. Biol. 13, 4276-4283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Waters H. L., Seetharam, S., Seidman, M. M. & Kraemer, K. H. (1993) J. Invest. Dermatol. 101, 744-748. [DOI] [PubMed] [Google Scholar]
- 9.Haracska L., Yu, S.-L., Johnson, R. E., Prakash, L. & Prakash, S. (2000) Nat. Genet. 25, 458-461. [DOI] [PubMed] [Google Scholar]
- 10.Haracska L., Prakash, S. & Prakash, L. (2000) Mol. Cell. Biol. 20, 8001-8007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Johnson R. E., Washington, M. T., Haracska, L., Prakash, S. & Prakash, L. (2000) Nature 406, 1015-1019. [DOI] [PubMed] [Google Scholar]
- 12.Johnson R. E., Haracska, L., Prakash, S. & Prakash, L. (2001) Mol. Cell. Biol. 21, 3558-3563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Haracska L., Unk, I., Johnson, R. E., Johansson, E., Burgers, P. M. J., Prakash, S. & Prakash, L. (2001) Genes Dev. 15, 945-954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Johnson R. E., Prakash, S. & Prakash, L. (2000) Proc. Natl. Acad. Sci. USA 97, 3838-3843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Suzuki N., Ohashi, E., Hayashi, K., Ohmori, H., Grollman, A. P. & Shibutani, S. (2001) Biochemistry 40, 15176-15183. [DOI] [PubMed] [Google Scholar]
- 16.Levine R. L., Miller, H., Grollman, A., Ohashi, E., Ohmori, H., Masutani, C., Hanaoka, F. & Moriya, M. (2001) J. Biol. Chem. 276, 18717-18721. [DOI] [PubMed] [Google Scholar]
- 17.Suzuki N., Ohashi, E., Kolbanovskiy, A., Geacintov, N. E., Grollman, A. P., Ohmori, H. & Shibutani, S. (2002) Biochemistry 41, 6100-6106. [DOI] [PubMed] [Google Scholar]
- 18.Washington M. T., Johnson, R. E., Prakash, L. & Prakash, S. (2002) Proc. Natl. Acad. Sci. USA 99, 1910-1914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lee S.-H., Kwong, A. D., Pan, Z.-Q. & Hurwitz, J. (1991) J. Biol. Chem. 266, 594-602. [PubMed] [Google Scholar]
- 20.Goodman M. F., Creighton, S., Bloom, L. B. & Petruska, J. (1993) Crit. Rev. Biochem. Mol. Biol. 28, 83-126. [DOI] [PubMed] [Google Scholar]
- 21.Mendelman L. V., Petruska, J. & Goodman, M. F. (1990) J. Biol. Chem. 265, 2338-2346. [PubMed] [Google Scholar]
- 22.Creighton S., Bloom, L. B. & Goodman, M. F. (1995) Methods Enzymol. 262, 232-256. [DOI] [PubMed] [Google Scholar]
- 23.Trincao J., Johnson, R. E., Escalante, C. R., Prakash, S., Prakash, L. & Aggarwal, A. K. (2001) Mol. Cell 8, 417-426. [DOI] [PubMed] [Google Scholar]