
Abstract
Calcium (Ca2+) homeostasis is fundamental for cell metabolism, proliferation, differentiation, and cell death. Elevation in intracellular Ca2+ concentration is dependent either on Ca2+ influx from the extracellular space through the plasma membrane, or on Ca2+ release from intracellular Ca2+ stores, such as the endoplasmic/sarcoplasmic reticulum (ER/SR). Mitochondria are also major components of calcium signalling, capable of modulating both the amplitude and the spatio-temporal patterns of Ca2+ signals. Recent studies revealed zones of close contact between the ER and mitochondria called MAMs (Mitochondria Associated Membranes) crucial for a correct communication between the two organelles, including the selective transmission of physiological and pathological Ca2+ signals from the ER to mitochondria. In this review, we summarize the most up-to-date findings on the modulation of intracellular Ca2+ release and Ca2+ uptake mechanisms. We also explore the tight interplay between ER- and mitochondria-mediated Ca2+ signalling, covering the structural and molecular properties of the zones of close contact between these two networks.
Similar content being viewed by others


Introduction
Increase of intracellular [Ca2+] can be elicited through two fundamental mechanisms: i) Ca2+ mobilization from intracellular stores, mainly the endoplasmic reticulum (ER), or ii) entry from the extracellular milieu through the opening of plasma membrane Ca2+ channels. Mitochondria are equally important in physiological Ca2+ signalling through a process unraveled by a series of works demonstrating that Ca2+ release from the ER results in cytosolic Ca2+ increases that are paralleled by similar or even larger cycles of mitochondrial calcium uptake, and subsequent release [1].
Mitochondrial Ca2+ accumulation is due to the large electrochemical gradient (mitochondrial membrane potential, ψmt, usually between -150 and -180 mV). Recent studies using electron tomography techniques revealed the presence of overlapping regions between ER and mitochondria separated by a minimum distance of 10-25 nm, that allows the direct physical association of ER proteins with components of the outer mitochondrial membrane (OMM) [2, 3]. These zones were identified as 'hotspots' and have pivotal roles in several cellular functions, including an highly efficient transmission of Ca2+ from the ER to the adjacent mitochondrial network that stimulates oxidative metabolism and, conversely, might enable the metabolically energized mitochondria to regulate ER Ca2+ homeostasis [4].
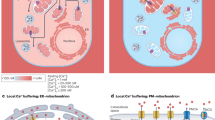
In this review, we attempt to give a condensed overview of the molecular aspects of intracellular calcium homeostasis, with special emphasis on the structural and molecular aspects of the ER (main intracellular Ca2+ stores), mitochondria (possessors of a series of complex systems for the influx/efflux of Ca2+) and MAMs (covering important roles in various cellular 'housekeeping' functions, including Ca2+ signalling) (Figure 1).

Representation of intracellular Ca2+ dynamics and MAMs proteins involved in ER-mitochondria Ca2+ cross-talk. A series of protein localized in MAMs (such as PML, AKT, grp-75, SIG-1R, Mfn-1/-2, BiP, AKT) regulate Ca2+ release from the ER and an efficient mitochondrial Ca2+ uptake, resulting in different functional outcomes. Cells generate Ca2+ signal through two mechanism that use internal and external sources of Ca2+. Calcium enters into the cell through channels and pumps situated on the plasma membrane; these are gated by voltage (VOCs) or external messengers (ROCs). A series of stimuli that act on cell surface receptors triggers the activation of PLC that catalyses the hydrolysis of phosphatidylinositol 4,5-biphosphate to IP3 and DAG. The binding of IP3 to its receptor IP3R stimulates ER Ca2+ release and consequently the transfer of Ca2+ (red dots) from ER to mitochondria. Mitochondrial surface directly interacts with the ER through contact sites defining hotspot Ca2+ signalling units. Mitochondrial Ca2+ import occurs through the mitochondrial Ca2+ uniporter (MCU) and the H+/Ca2+ exchanger LETM1; conversely, NCLX, mitochondrial Na+/Ca2+ exchanger, together with the PTP, export Ca2+ from the matrix. Ca2+ levels return to resting conditions through a series of channels and pumps: PMCA and NCX permit the ion extrusion into the extracellular milieu, SERCA (situated on the ER) and SPCA (on the Golgi apparatus) re-establish basal Ca2+ levels in intracellular stores. Abbreviations: ANT, adenosine nucleoside transporter; ETC, electron transport chain; HK, hexokinase; CD, cyclophilin D; CK, creatine kinase; BR, benzodiazepine receptor.
1. Mitochondrial structure
Mitochondria, considered the "biochemical powerhouse" of the cell, are dynamic and plastic organelles, constantly subjected to remodeling, involved in a number of crucial metabolic roles, such as the tricarboxylic acid (TCA) cycle and β-oxidation of fatty acids [5]. These organelles possess two membranes that give rise to different functional regions: the inner and outer mitochondrial membranes themselves (IMM and OMM, respectively), the cytosolic side of the OMM, the intermembrane space (IMS), and the matrix. This somewhat unusual composition enables the occurrence of a wide variety of reactions, including oxidative phosphorylation. The IMM harbours complexes of the respiratory chain, ATP synthase, and enzymes involved in heme biosynthesis [2, 6]. The OMM is also very rich in proteins. Small molecules (< 10 kDa) can pass freely between the cytoplasm and the IMS due to the presence of a number of porins. Conversely, the IMM is completely impermeable even to small molecules, including protons (with the exception of O2, CO2, and H2O). This peculiarity enables the complexes of the respiratory chain to build up a proton gradient across the IMM, required for oxidative phosphorylation. The resulting electrochemical proton gradient forms the basis of the ψmt and is utilized to generate chemical energy in the form of ATP. This ψmt is also one of the crucial factors responsible for Ca2+ uptake. Ca2+ can freely cross the OMM but in order to enter the matrix it must pass through the mitochondrial Ca2+uniporter (MCU), an IMM-located channel driven by a large electrochemical gradient [7–10].
2. ER structure
The endoplasmic reticulum (ER) is considered the largest individual intracellular organelle. It consists of a three-dimensional network of endomembranes, constituting a complex grid of microtubules and cisternae. The domains accounting for the ER are functionally and structurally distinct. Historically, ER histology describes three types of ER: smooth ER, rough ER and the nuclear envelope (NE) [11]. This division demonstrates the individual role of each particular ER type. A morphological characterization describes the NE and peripheral ER, the later being a network of tubules and sheets reaching the most remote areas of the cell. Such structural diversity of the ER, reviewed extensively by a number of authors [12–15], is related to the variety of cellular functions played by the organelle. For such a large organelle, the ER is unexpectedly plastic [16]. This plasticity is a crucial morphological characteristic of the ER and the remodeling ability seems to be correlated with the diversity of its functions. The most important activities of ER regard protein synthesis and maturation. Protein synthesis occurs in ribosome-rich rough ER, whereas their post-translational processing is carried out by an extensive group of chaperone proteins [17]. Importantly, the ER acts as a transport route enabling the delivery of a number of proteins to their destination [18]. The third function is that the ER is a dynamic reservoir of Ca2+ ions, which can be activated by both electrical and chemical cell stimulation [16, 19]. This feature renders this organelle an indispensable source of Ca2+ in many aspects of physiological signalling.
3. MAM structure
The association between the ER and mitochondria was first described by Copeland and Dalton over 50 years ago in pseudobranch gland cells [20]. However, it was only in the beginning of the 1970s that mitochondria-ER contacts were visualized by a number of research groups [21, 22]. Further developments in microscopy techniques provided researchers with the ability to perform detailed analyses with high resolution in 3 dimensions [23, 24]. The interactions between these organelles at the contact places are so tight and strong that subcellular fractionation enabled the performance of a step through which a unique fraction, originally named as 'mitochondria-associated membranes" (MAMs fraction), can be isolated [25]. Further developments in experimental procedures enabled scientists to isolate pure MAM fractions from yeast, different organs and tissues, as well as various cell lines [25, 26].
4. The intracellular Ca2+-signalling network
Intracellular Ca2+ signalling is versatile and fundamental for the regulation of multiple cellular processes, including development, proliferation, secretion, gene activation and cell death. The universality of Ca2+ as an intracellular messenger depends on its enormous versatility in terms of speed, amplitude, and spatio-temporal patterning. The Ca2+-signalling network can be divided into four functional units:
-
signalling is triggered by a stimulus that generates various Ca2+ -mobilizing signals;
-
the latter activate the ON mechanisms that feed Ca2+ into the cytoplasm;
-
Ca2+ functions as a messenger to stimulate numerous Ca2+-sensitive processes; and finally,
-
the OFF mechanisms, composed of pumps and exchangers, remove Ca2+ from the cytoplasm to restore the resting state [27].
Inside cells, Ca2+ concentration ([Ca2+]) is controlled by the simultaneous interplay of multiple counteracting processes, which can be split into "ON mechanisms" and "OFF mechanisms". With these, cells maintain a rigid control over the cytosolic level of Ca2+. In resting conditions, cells maintain a low Ca2+ concentration in the cytoplasm (around 100 nM). Ca2+ influx from the extracellular space (that posses a concentration around 1-2 mM), or Ca2+ release from intracellular Ca2+ stores, such as the ER (with concentrations of 250-600 μM) generate the intracellular calcium signals [28].
5. The Ca2+-induced Ca2+ release process
Cells generate their Ca2+ signals through two fundamental mechanisms that make use of internal and external sources of calcium. Ca2+-mobilizing signals from internal stores are generated by stimuli that act through a variety of cell-surface receptors, such as G-protein-linked receptors and receptor tyrosine kinases. When a ligand binds its specific receptor in the plasma membrane, the occupied receptor causes GDP-GTP exchange on an associated G-protein, Gq, that activates a specific membrane-bound phospholipase C (PLC), which in turn catalyzes the production of the two second messengers, diacylclycerol (DAG) and inositol-1,4,5-trisphosphate (IP3), by hydrolysis of phosphatidylinositol 4,5-bisphosphate in the plasma membrane. IP3, a water-soluble compound, diffuses from the plasma membrane to the endoplasmic reticulum, where it binds to specific IP3 receptors (IP3Rs) and causes Ca2+ channels within the ER to open. Sequestered Ca2+ is thus released into the cytosol, and the cytosolic [Ca2+] rises sharply to about 10 μM [29].
On the plasma membrane there is a large family of Ca2+ entry channels, important for the "ON reaction", which can be defined by the way in which they are activated. The most known are voltage-operated channels (VOCs) that are triggered by membrane depolarization. Other channels, called receptor-operated channels (ROCs), are sensitive to the binding of different external signals, usually transmitters. Finally, store-operated channels (SOCs), respond to the depletion of internal Ca2+ stores and contribute to the spatio-temporal pattern of Ca2+ waves [30].
As mentioned above, IP3Rs are the most important actors in Ca2+ release from internal stores, controlled by Ca2+ itself that acts either on the luminal or cytoplasmic sides of the channel.
5.1 IP3Rs
IP3Rs, the main Ca2+-release channels in the ER of most cell types, consist of four subunits of about 310 kDa each with a similar general structure. In mammals, three different genes encode for three different isoforms (IP3R1, -2 and -3). Structurally, the proteins have a cytoplasmic N-terminal hydrophobic region, predicted to contain six membrane-spanning helices, and a relatively short cytoplasmic C-terminus. Functionally, the N-terminal domain contains the IP3-binding domain and a "regulatory"/"coupling" domain. IP3Rs can be modulated primarily by IP3 and Ca2+ itself; the latter can also regulate IP3Rs indirectly through calmodulin (CaM); other modulators include phosphorylation by Ca2+/CaM-dependent kinase II (CaMKII), cGMP-dependent protein kinase (PKG), protein kinase C (PKC), and cAMP-dependent protein kinase (protein kinase A, PKA). This suggests that the IP3R works as a crosstalk station between Ca2+ signalling and phosphorylation [31]. As a confirmation of this, IP3R isoforms contain multiple phosphorylation consensus sites and many docking sites for protein kinases and phosphatases, and at least 15 different protein kinases are known to directly phosphorylate IP3R [32].
6. Mitochondria in calcium signalling
Mitochondria are important components of the "OFF" reaction since they modulate both the amplitude [33] and the spatio-temporal patterns of Ca2+ signals [34, 35].
That mitochondria can accumulate certain ions from the suspending medium was first observed in the early 1960s, when was discovered that isolated mitochondria from rat liver, kidney, brain and heart can accumulate large net amounts of Ca2+ from the suspending medium during electron transport, up to several hundred times the initial Ca2+ content [36].
In these studies, initial velocities of energy-dependent Ca2+ uptake were measured by stopped-flow and dual-wavelength techniques in mitochondria isolated from hearts of rats. The first rate of Ca2+ uptake shows that the initial velocity of Ca2+ uptake was slow at low concentrations of Ca2+ and increased sigmoidally to 10 nM Ca2+/s/mg protein at 300 μM Ca2+. Similar results were obtained by the employment of mitochondria subjected of a wide range of mitochondrial protein in the medium (0.5-10 mg/ml), when these organelles were oxidizing glutamate-malate and when acetate was replacing phosphate as a permanent anion [37].
Comparable rates of Ca2+ uptake and sigmoidal plots were obtained with mitochondria from other mammalian hearts, as like guinea pigs, squirrels, pigeons, and frogs where the rate of Ca2+ uptake was 0.05 nM/mg/s at 5 μM Ca2+ and increased sigmoidally to 8 nM/mg/s at 200 μM Ca2+[38].
Mitochondrial Ca2+ uptake plays a key role in the regulation of many cells functions, ranging from ATP production to cell death. Increases in mitochondrial calcium activates several dehydrogenases and carriers, inducing an increase in the respiratory rate, H+ extrusion, and ATP production necessary for the correct energy state of the cell. However, prolonged increase in [Ca2+]m leads to the opening of the mitochondrial permeability transition pore (PTP), a critical event driving to cell death by apoptosis [39].
Although it is generally accepted that cellular energy metabolism, survival and death are controlled by mitochondrial calcium signals, the underlying molecular mechanisms have been completely elucidated yet. Several studies have identified three essential proteins mediating the processes of calcium influx and efflux.
6.1 Mitochondrial Calcium Uniporter (MCU)
The main transporters involved in the uptake of Ca2+ into mitochondria is the MCU, characterized by a low affinity for Ca2+; in fact, MCU takes up Ca2+ in the micromolar range and experiments in permeabilized cells report a K d of the uniporter of 10 μM [40]. In addition, a biphasic effect of calcium on the MCU has been reported: beyond a certain level, cytosolic Ca2+ inactivates the uniporter, preventing further Ca2+ uptake and this process might avoid an excessive accumulation of the cation in mitochondria [41].
In spite of repeated efforts by different researchers, the molecular identity of the MCU has remained elusive. Among the early candidates proposed for the MCU were the uncoupling proteins UCP2/3 [42], but experiments in different tissues of mice lacking UCP2 and UCP3 showed a normal Ca2+ uptake [43]. Recently, Perocchi and colleagues [44] demonstrated that MICU1 (mitochondrial calcium uptake 1), also known as FLJ12684 or CBARA1, has a key role in regulating the classically defined uniporter. MICU1 is associated with the IMM and has two canonical EF hands that are essential for its activity and it caused a significant suppression of the [Ca2+]m signal evoked by an IP3-linked agonist. Silencing MICU1 does not impair mitochondrial respiration or membrane potential but abolishes Ca2+ entry in intact and permeabilized cells, and attenuates the metabolic coupling between cytosolic Ca2+ transients and activation of matrix dehydrogenases.
More recently, in 2011, two distinct laboratories have been identified a transmembrane protein (CCDC109A) that fulfilling the criteria for being the MCU [9, 10]. Indeed, in planar lipid bilayers CCDC109A showed channel activity with electrophysiological properties as those previously reported for the MCU [8]. The over-expression of CCDC109A (that now is called "MCU"), increases mitochondrial Ca2+ uptake and sensitizes cells to apoptotic stimuli, and the employment of short interfering RNA (siRNA) silencing of MCU strongly reduced mitochondrial Ca2+ uptake. This reduction is specific for mitochondria (Ca2+ cytosolic levels remain almost unaffected), does not induce impairment of the electrochemical gradient or change in mitochondrial morphology and the induction of specific mutations at the level of the putative pore-forming region reduce the mitochondrial calcium uptake and blocks the channel activity of the protein [9, 10].
To conclude, MCU and MICU1 are critical for the correct mitochondrial calcium uptake: the first one can be considered the main component of the uniporter, while MICU1 as a fundamental regulator.
6.2 LETM1
As described above, MCU only takes up Ca2+ in the micromolar range, but evidence has shown that mitochondria are able to take up Ca2+ also at much lower concentrations, as recently reported by Jang and colleagues who identified a high-affinity mitochondrial Ca2+/H+ exchanger capable of importing calcium in the nanomolar range [45]. This group conducted a genome-wide RNAi screen in Drosophila cells stably expressing a mitochondria-targeted ratiometric Pericam and identified the gene CG4589 (Drosophila homolog of the human gene LETM1, leucine zipper-EF-hand containing transmembrane protein 1) as a regulator of mitochondrial Ca2+ and H+ concentrations, supporting electrogenic import of Ca2+ (one Ca2+ in for one H+ out).
However, the effective role of LETM1 as Ca2+/H+ exchanger still remains a subject of discussion, since its activity is blocked by treatment with CGP37157 (channel inhibitor that mediates mitochondrial calcium efflux) and red/Ru360 (inhibitor of MCU). Furthermore, LETM1 is associated with K+ homeostasis, and the loss of LETM1 lowers mitochondrial membrane potential, and the mitochondrial H+/Ca2+ exchanger turned out to be non-electrogenic (one Ca2+ in for two H+ out) [45, 46].
6.3 NCLX/NCKX6
A Na+-dependent mechanism that mediates mitochondrial Ca2+ efflux has been demonstrated, but the molecular identity of this transporter has also remained elusive. In a recent study, Palty and co-workers showed that the Na+/Ca2+ exchanger NCLX is enriched in mitochondria, where it is localized to the cristae [47]. This protein was identified as a member of the Na+/Ca2+ exchanger situated in the ER or plasma membrane, but Palty et al., shown that in several tissues endogenous NCLX is enriched primarily in mitochondria, but not in ER and plasma membrane. The same observation is achieved overexpressing the protein in different cell lines, and the results show that expression of NCLX enhances mitochondrial Ca2+ efflux; this is blocked by CGP37157 and by mutations in the catalytic site of NCLX. Besides, the role of NCLX as a mitochondrial Na+/Ca2+ exchanger is supported by evidence that NCLX mediates Li+/Ca2+ exchange, a functional property that, among NCX proteins, is shared exclusively with the mitochondrial exchanger [47].
7. Intracellular calcium extrusion mechanism
Once it activates its downstream targets, Ca2+ has carried out its functions and needs to be rapidly removed from the cytosol to restore the resting levels of approximately 100 nM. For this purpose, the cell uses the combined activity of Ca2+ extrusion mechanisms (such as PMCA and NCX) and mechanisms that refill the intracellular stores (like sarco-endoplasmic reticulum Ca2+-ATPases, SERCAs, and the secretory-pathway Ca2+-ATPases, SPCAs, of the Golgi apparatus).
7.1 PMCA
The plasma membrane calcium ATPases (PMCA) is localized on the plasma membrane and couples ATP hydrolysis to the maintenance of appropriate cytoplasmic calcium levels by removing calcium from the cytosol to the extracellular spaces. There are at least four different PMCA isoforms (PMCA1-4) and several splice variants (about 26) that are encoded by four independent genes. Some of these are ubiquitously expressed in the organism (PMCA1 and -4), while others (PMCA2 and -3) have a tissue-specific expression patterns. Structurally, PMCAs consist of 10 transmembrane domains, two major intracellular loops, and N- and C-cytoplasmic domains. The pump operates with high Ca2+ affinity and low transport capacity, with a 1:1 Ca2+/ATP stoichiometry. Under optimal conditions, the K d of PMCA for Ca2+ is about 10-30 μM in resting conditions and about 0.2-0.5 μM in activated conditions [48].
It has been demonstrated that PMCA operates as a Ca2+/H+ exchanger and, even if the exact stoichiometry is not well defined, a recent study suggests that the Ca2+:H+ ratio is 1:2 and that the activity of the pump is insensitive to variations of membrane potential [49].
7.2 NCX
NCX (Na2+/Ca2+ exchanger) is a plasma-membrane enzyme, mainly located in excitable tissue, which carries out the efflux of one Ca2+ against an influx of 3 Na+. NCX easily reverses its direction and brings Ca2+ into the cells if the Na+ concentration gradient decreases and/or the membrane potential becomes less negative. Three distinct genes are known to encode as many isoforms, namely NCX1, NCX2 and NCX3 [50]. The first one has an ubiquitous distribution, while NCX2 is expressed primarily in the brain, and NCX3 in the skeletal muscle. The NCX1 protein contains 11 putative transmembrane domains, divided into two sets of putative transmembrane domains separated by a large intracellular loop mainly responsible for the transport of the Na+ and Ca2+ across the membrane [29].
7.3 SPCA
SPCAs are the newest addition to the family of phosphorylation-type ATPases and they are responsible for supplying the lumen of the Golgi apparatus with Ca2+. Unlike other Ca2+-ATPase pumps, SPCA pumps are not electrogenic; in fact, they do not counter-transport H+ to the outside since protons are essential for the correct development and functioning of the Golgi vesicle [51]. In addition to Ca2+ transport, the most important property of SPCA pumps is also to transport efficiently Mn2+ into the Golgi, as this is a necessary cation for enzymes present in the lumen of the Golgi compartment. SPCAs function through a reversibly cycle between an E1- and an E2-conformation. In the cytosol, the high-affinity binding site of the protein in E1-conformation binds Ca2+ (or Mn2+), and phosphorylation by ATP creates an high-energy phosphoenzyme intermediate. This enzyme undergoes a rate-limiting transition to the lower-energy state, E2, and simultaneously Ca2+ (or Mn2+) moves through the transmembrane pore and is released into the lumen of the Golgi apparatus. As a last step, the protein returns to the dephosphorylated state [52].
7.4 SERCA
SERCA is a pump identified in 1961-1962 in a skeletal muscle fraction. It is localized in the membranes of endo(sarco)plasmic reticulum and couples ATP hydrolysis to the transport of Ca2+ from cytoplasm to lumen. Early studies shown that the pump counter-transported H+ in exchange for two Ca2+ per ATP hydrolyzed. However, it has been noticed that fewer than four H+ were released to the cytosol per two Ca2+ pumped, showing that the transport reaction was only partly electrogenic [53]. Like other Ca2+-ATPase pumps, SERCAs exist in two conformational states. The E1 on the cytosolic site, in which the enzyme has high Ca2+ affinity, and the E2 state, in which the lower Ca2+ affinity leads to the release of Ca2+ on the opposite side. This cycle has a number of other states that occur upon binding of Ca2+, involving a series of structural changes in the cytoplasmic sector and in the transmembrane domain, necessary for completing the catalytic cycle. The peculiarity of SERCAs in respect to the other Ca2+-ATPase pumps is to have two Ca2+ binding sites, enabling the existence of a Ca2+/ATP transport stoichiometry of 2.0 [54].
8. MAMs, a functional link between ER and mitochondria
As described above, mitochondria and endoplasmic reticulum networks are fundamental for the maintenance of calcium homeostasis. Recently, different studies have documented the crucial role that MAMs play in intracellular Ca2+ signalling. The physical proximity of the ER to mitochondria enables a direct, selective transmission of physiological and pathological Ca2+ signals [1], an aspect highly variable between cell types. In fact, mitochondria are not always morphologically continuous, functionally homogenous and associated to ER. At demonstration of this, different works revealed the existence of a largely interconnected mitochondria network akin to ER in HeLa cells, COS-7 cells, cardiac myocytes and rat hepatocytes [24, 55]. Contrary, it has been also reported that mitochondria can exist as two distinct populations, one in perinuclear position and the other one in cell periphery, with different biochemical and respiratory properties [56]. Or, again, mitochondria within individual cells are morphologically heterogeneous and appear as distinct entities [57]. These different aspects could carry out diverse aspect of mitochondrial functions, in particular Ca2+ sequestration, fundamental for the regulation of mitochondrial metabolism and regulation of apoptosis [39]. Lately, it has been demonstrated that the juxtaposition between ER and mitochondria is also regulated by cellular status. In fact, a condition of starvation (an autophagic trigger) leads to PKA activation, which in turn phosphorylates the pro-fission dynamin-related protein 1 (DRP1) with consequent mitochondria elongation in a network of highly interconnected organelles. This mitochondrial elongation protects cells from death and is required to sustain ATP levels and viability [58].
The MAMs, these ER-contiguous membranes, contain multiple phospholipid- and glycosphingolipid-synthesizing enzymes, including long-chain fatty acid-CoA ligase type 4 (FACL4) and phosphatidylserine synthase-1 (PSS-1), enzymes required for cholesterol and ceramide biosynthesis, enzymes involved in glucose metabolism and support direct transfer of lipids between the ER and mitochondria [1, 59]. MAMs also constitute a calcium signaling hub regulating ER chaperone-assisted folding of newly synthesized proteins, the mitochondria-localized dehydrogenases activity, and the activation of Ca2+-dependent enzymes that execute cell death programs [60]. The importance of MAMs began to emerge when it was found that, after cell stimulation, mitochondria were able to uptake Ca2+ directly from IP3Rs [8, 61]. It was thus possible to identify specialized signalling microdomains, selectively enriched in critical Ca2+ signalling elements, labelled as "hotspots zones", where Ca2+ is transferred from the ER into mitochondria (Table 1). The interactions between the two organelles are modulated by mitochondria-shaping proteins and chaperone proteins. MFN-1 and -2 (mitofusin-1 and -2) belong to the first group and stabilize the interaction between adjacent mitochondria, regulate ER morphology and calcium homeostasis, and directly tether ER to mitochondria, thus facilitating efficient Ca2+ uptake by mitochondria [23]. The hsp70 homologous cytosolic chaperone grp75 (glucose-regulated protein 75) tethers the N-terminal domain of the type-1 IP3Rs to the isoform 1 of VDAC, generating a molecular bridge that enhances the Ca2+ accumulation in mitochondria [62].
Recently identified, the Sigma-1 ER receptor (Sig-1R) selectively resides at the MAMs, forms a Ca2+-sensitive chaperone complex with BiP/GRP78 (78-kDa glucose-regulated protein GRP78, also referred to as the immunoglobulin binding protein BiP) and associates with isoform 3 of IP3R. Upon activation of IP3Rs, which causes the decrease of Ca2+ concentration at the MAM, redistribution of Sig-1Rs occurs, from MAMs to the periphery of the ER: here Sig-1Rs dissociates from BiP/GRP78 and the chaperone activity of free Sig-1Rs attenuates the aggregation of IP3R3 [63].
Obviously, other proteins that are required to modulate calcium mobilization upon cellular stimulation are directed to MAMs. An example is the anti-apoptotic protein AKT/PKB that, in response to survival signals, is recruited to MAMs in order to inactivate IP3R3, significantly reducing ER-Ca2+ release activity with a diminished cellular sensitivity to apoptotic stimuli [64]. In turn, this event determines the PML (promyelocytic leukemia protein)-mediated recruitment of phosphatase PP2a (protein phosphatase 2a) at the MAMs to switch off the kinase. Also cytochrome c, which is released from mitochondria upon activation of apoptotic pathways, can bind IP3Rs at the MAMs, further activating the Ca2+ flux and enhancing apoptotic signaling [65]. Recently, two interesting proteins with a marked regulatory effect on cell survival through changes in Ca2+ have been identified in the zones of mitochondria-ER association.
p66Shc (a 66-kDa isoform of the growth factor adapter Shc) is a cytosolic adaptor protein, profoundly involved in the cellular response to oxidative stress. This protein has also been found in the MAM fraction. Its direct relation to mitochondrial ROS production has been repeatedly documented, also by our groups [66–68]. We found that the level of p66Shc in the MAM fraction is age-dependent and corresponds well to the mitochondrial ROS production which is found to increase with age [69].
PML is another protein recently identified in the MAM fraction. There, it is believed to regulate the ER machinery responsible for Ca2+ release. The lack of PML results in a decreased Ca2+ release from the ER and a subsequent lower Ca2+ influx into mitochondria. Detailed studies on the role of PML protein in the MAM fraction contributed to the formation of the hypothesis that this protein regulates cell survival through the ER-cytosol/mitochondria calcium signalling [70] (Figure 2).

PML and p66Shc regulates cell span at the MAMs level. The tumor suppressor PML in resting conditions resides in a specific multi-protein complex with IP3R, PP2a and AKT, essential for a normal Ca2+ flux from ER to mitochondria and, consequently, for correct apoptosis levels (upper panel). Aging and ROS determine phosphorylation and accumulation of p66Shc in the MAMs fraction. The presence of phospho-p66Shc at the mitochondrial level determines alterations in mitochondrial homeostasis, including Ca2+ signalling, and ultimately increases apoptotic and senescence responses (lower panel).
Conclusion
In this review, we have sketched out the main features of the intracellular Ca2+-signalling toolkit and the elaborate relationship between ER and mitochondria. It has been demonstrated that the physical interactions between ER and mitochondria, known as MAMs, are essential for functions of the two organelles, and that these also enable an highly efficient transmission of Ca2+ from the ER to mitochondria. In this regard, it becomes evident that the ER-mitochondria interface points deeply affect intracellular Ca2+ signalling and are fundamental for different functional outcomes, such as cell metabolism or induction of cell death.
Abbreviations
- ψmt:
-
mitochondrial membrane potential
- AKT/PKB:
-
Protein kinase B
- BiP/GRP78:
-
Binding immunoglobulin Protein/78-kDa glucose-regulated protein
- Ca2+:
-
calcium ions
- [Ca2+]:
-
Ca2+ concentration
- [Ca2+]c:
-
cytosolic Ca2+ concentration
- [Ca2+]m:
-
mitochondrial Ca2+ concentration
- CaM:
-
calmodulin
- CaMKII:
-
calmodulin-dependent protein kinase II
- DAG:
-
diacylclycerol
- DRP1:
-
dynamin-related protein 1
- ER:
-
endoplasmic reticulum
- FACL4:
-
long-chain fatty acid-CoA ligase type 4
- grp75:
-
glucose-regulated protein 75
- IMM:
-
inner mitochondrial membrane
- IMS:
-
intermembrane space
- IP3:
-
inositol 1,4,5-trisphosphate
- IP3R:
-
inositol 1,4,5-trisphosphate receptor
- Letm1:
-
leucine zipper-EF-hand containing transmembrane protein 1
- MAMs:
-
mitochondria-associated membranes
- MCU:
-
mitochondrial Ca2+ uniporter
- MICU1:
-
mitochondrial calcium uptake 1
- MFN-1/-2:
-
mitofusin-1: -2
- NCX:
-
Na2+/Ca2+ exchanger
- NE:
-
nuclear envelope
- OMM:
-
outer mitochondrial membrane
- p66Shc:
-
66-kDa isoform of the growth factor adapter shc
- PKA:
-
protein kinase A
- PKC:
-
protein kinase C
- PKG:
-
cGMP-dependent protein kinase
- PLC:
-
phospholipase C
- PMCA:
-
plasma membrane Ca2+ ATPase
- PML:
-
promyelocytic leukemia protein
- PP2a:
-
protein phosphatase 2a
- PSS-1:
-
phosphatidylserine synthase-1
- PTP:
-
permeability transition pore
- ROCs:
-
receptor operated Ca2+ channels
- ROS:
-
reactive oxygen species
- SERCA:
-
sarco-endoplasmic reticulum Ca2+ ATPase
- Sig-1R:
-
Sigma-1 receptor
- SMOCs:
-
second messenger operated Ca2+ channels
- SPCA:
-
secretory-pathway Ca2+-ATPase
- SR:
-
sarcoplasmic reticulum
- UCP:
-
uncoupling protein
- VDAC:
-
voltage-dependent anion channel
- VOCs:
-
voltage operated Ca2+ channels.
References
Giorgi C, De Stefani D, Bononi A, Rizzuto R, Pinton P: Structural and functional link between the mitochondrial network and the endoplasmic reticulum. Int J Biochem Cell Biol. 2009, 41 (10): 1817-27. 10.1016/j.biocel.2009.04.010.
Fernie AR, Carrari F, Sweetlove LJ: Respiratory metabolism: glycolysis, the TCA cycle and mitochondrial electron transport. Curr Opin Plant Biol. 2004, 7 (3): 254-61. 10.1016/j.pbi.2004.03.007.
Rizzuto R, Duchen MR, Pozzan T: Flirting in little space: the ER/mitochondria Ca2+ liaison. Sci STKE. 2004, 2004 (215): re1-
Csordás G, Renken C, Várnai P, Walter L, Weaver D, Buttle KF, Balla T, Mannella CA, Hajnóczky G: Structural and functional features and significance of the physical linkage between ER and mitochondria. J Cell Biol. 2006, 174 (7): 915-21. 10.1083/jcb.200604016.
McBride HM, Neuspiel M, Wasiak S: Mitochondria: more than just a powerhouse. Curr Biol. 2006, 16 (14): R551-60. 10.1016/j.cub.2006.06.054.
Rich PR: The molecular machinery of Keilin's respiratory chain. Biochem Soc Trans. 2003, 31 (Pt 6): 1095-105.
Nicholls DG, Crompton M: Mitochondrial calcium transport. FEBS Lett. 1980, 111 (2): 261-8. 10.1016/0014-5793(80)80806-4.
Kirichok Y, Krapivinsky G, Clapham DE: The mitochondrial calcium uniporter is a highly selective ion channel. Nature. 2004, 427 (6972): 360-4. 10.1038/nature02246.
De Stefani D, Raffaello A, Teardo E, Szabò I, Rizzuto R: A forty-kilodalton protein of the inner membrane is the mitochondrial calcium uniporter. Nature. 2011
Baughman JM, Perocchi F, Girgis HS, Plovanich M, Belcher-Timme CA, Sancak Y, Bao XR, Strittmatter L, Goldberger O, Bogorad RL, Koteliansky V, Mootha VK: Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature. 2011
Baumann O, Walz B: Endoplasmic reticulum of animal cells and its organization into structural and functional domains. Int Rev Cytol. 2001, 205: 149-214.
Voeltz GK, Prinz WA, Shibata Y, Rist JM, Rapoport TA: A class of membrane proteins shaping the tubular endoplasmic reticulum. Cell. 2006, 124 (3): 573-86. 10.1016/j.cell.2005.11.047.
Shibata Y, Voeltz GK, Rapoport TA: Rough sheets and smooth tubules. Cell. 2006, 126 (3): 435-9. 10.1016/j.cell.2006.07.019.
English AR, Zurek N, Voeltz GK: Peripheral ER structure and function. Curr Opin Cell Biol. 2009, 21 (4): 596-602. 10.1016/j.ceb.2009.04.004.
Shibata Y, Shemesh T, Prinz WA, Palazzo AF, Kozlov MM, Rapoport TA: Mechanisms determining the morphology of the peripheral ER. Cell. 2010, 143 (5): 774-88. 10.1016/j.cell.2010.11.007.
Bootman MD, Petersen OH, Verkhratsky A: The endoplasmic reticulum is a focal point for co-ordination of cellular activity. Cell Calcium. 2002, 32 (5-6): 231-4. 10.1016/S0143416002002002.
Chevet E, Cameron PH, Pelletier MF, Thomas DY, Bergeron JJ: The endoplasmic reticulum: integration of protein folding, quality control, signaling and degradation. Curr Opin Struct Biol. 2001, 11 (1): 120-4. 10.1016/S0959-440X(00)00168-8.
Palade G: Intracellular aspects of the process of protein synthesis. Science. 1975, 189 (4200): 347-58. 10.1126/science.1096303.
Verkhratsky A, Petersen OH: The endoplasmic reticulum as an integrating signalling organelle: from neuronal signalling to neuronal death. Eur J Pharmacol. 2002, 447 (2-3): 141-54. 10.1016/S0014-2999(02)01838-1.
Copeland DE, Dalton AJ: An association between mitochondria and the endoplasmic reticulum in cells of the pseudobranch gland of a teleost. J Biophys Biochem Cytol. 1959, 5 (3): 393-6. 10.1083/jcb.5.3.393.
Lewis JA, Tata JR: A rapidly sedimenting fraction of rat liver endoplasmic reticulum. J Cell Sci. 1973, 13 (2): 447-59.
Morre DJ, Merritt WD, Lembi CA: Connections between mitochondria and endoplasmic reticulum in rat liver and onion stem. Protoplasma. 1971, 73 (1): 43-9. 10.1007/BF01286410.
de Brito OM, Scorrano L: Mitofusin 2 tethers endoplasmic reticulum to mitochondria. Nature. 2008, 456 (7222): 605-10. 10.1038/nature07534.
Rizzuto R, Pinton P, Carrington W, Fay FS, Fogarty KE, Lifshitz LM, Tuft RA, Pozzan T: Close contacts with the endoplasmic reticulum as determinants of mitochondrial Ca2+ responses. Science. 1998, 280 (5370): 1763-6. 10.1126/science.280.5370.1763.
Vance JE, Stone SJ, Faust JR: Abnormalities in mitochondria-associated membranes and phospholipid biosynthetic enzymes in the mnd/mnd mouse model of neuronal ceroid lipofuscinosis. Biochim Biophys Acta. 1997, 1344 (3): 286-99.
Wieckowski MR, Giorgi C, Lebiedzinska M, Duszynski J, Pinton P: Isolation of mitochondria-associated membranes and mitochondria from animal tissues and cells. Nat Protoc. 2009, 4 (11): 1582-90. 10.1038/nprot.2009.151.
Berridge MJ, Lipp P, Bootman MD: The versatility and universality of calcium signalling. Nat Rev Mol Cell Biol. 2000, 1 (1): 11-21.
Demaurex N, Frieden M: Measurements of the free luminal ER Ca2+ concentration with targeted "cameleon" fluorescent proteins. Cell Calcium. 2003, 34 (2): 109-19. 10.1016/S0143-4160(03)00081-2.
Berridge MJ, Bootman MD, Roderick HL: Calcium signalling: dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol. 2003, 4 (7): 517-29. 10.1038/nrm1155.
Balla T: Regulation of Ca2+ entry by inositol lipids in mammalian cells by multiple mechanisms. Cell Calcium. 2009, 45 (6): 527-34. 10.1016/j.ceca.2009.03.013.
Carafoli E, Santella L, Branca D, Brini M: Generation, control, and processing of cellular calcium signals. Crit Rev Biochem Mol Biol. 2001, 36 (2): 107-260. 10.1080/20014091074183.
Vanderheyden V, Devogelaere B, Missiaen L, De Smedt H, Bultynck G, Parys JB: Regulation of inositol 1,4,5-trisphosphate-induced Ca2+ release by reversible phosphorylation and dephosphorylation. Biochim Biophys Acta. 2009, 1793 (6): 959-70. 10.1016/j.bbamcr.2008.12.003.
Budd SL, Nicholls DG: A reevaluation of the role of mitochondria in neuronal Ca2+ homeostasis. J Neurochem. 1996, 66 (1): 403-11.
Duchen MR: Contributions of mitochondria to animal physiology: from homeostatic sensor to calcium signalling and cell death. J Physiol. 1999, 516 (Pt 1): 1-17.
Jouaville LS, Ichas F, Holmuhamedov EL, Camacho P, Lechleiter JD: Synchronization of calcium waves by mitochondrial substrates in Xenopus laevis oocytes. Nature. 1995, 377 (6548): 438-41. 10.1038/377438a0.
Rossi CS, Lehninger AL: Stoichiometric relationships between mitochondrialion accumulation and oxidative phosphorylation. Biochem Biophys Res Commun. 1963, 11: 441-6. 10.1016/0006-291X(63)90089-5.
Scarpa A, Graziotti P: Mechanisms for intracellular calcium regulation in heart. I. Stopped-flow measurements of Ca2+ uptake by cardiac mitochondria. J Gen Physiol. 1973, 62 (6): 756-72. 10.1085/jgp.62.6.756.
Lehninger AL: Mitochondria and calcium ion transport. Biochem J. 1970, 119 (2): 129-38.
Giorgi C, Romagnoli A, Pinton P, Rizzuto R: Ca2+ signaling, mitochondria and cell death. Curr Mol Med. 2008, 8 (2): 119-30. 10.2174/156652408783769571.
Bragadin M, Pozzan T, Azzone GF: Kinetics of Ca2+ carrier in rat liver mitochondria. Biochemistry. 1979, 18 (26): 5972-8. 10.1021/bi00593a033.
Moreau B, Nelson C, Parekh AB: Biphasic regulation of mitochondrial Ca2+ uptake by cytosolic Ca2+ concentration. Curr Biol. 2006, 16 (16): 1672-7. 10.1016/j.cub.2006.06.059.
Trenker M, Malli R, Fertschai I, Levak-Frank S, Graier WF: Uncoupling proteins 2 and 3 are fundamental for mitochondrial Ca2+ uniport. Nat Cell Biol. 2007, 9 (4): 445-52. 10.1038/ncb1556.
Brookes PS, Parker N, Buckingham JA, Vidal-Puig A, Halestrap AP, Gunter TE, Nicholls DG, Bernardi P, Lemasters JJ, Brand MD: UCPs--unlikely calcium porters. Nat Cell Biol. 2008, 10 (11): 1235-7. 10.1038/ncb1108-1235. author reply 1237-40
Perocchi F, Gohil VM, Girgis HS, Bao XR, McCombs JE, Palmer AE, Mootha VK: MICU1 encodes a mitochondrial EF hand protein required for Ca2+ uptake. Nature. 2010, 467 (7313): 291-6. 10.1038/nature09358.
Jiang D, Zhao L, Clapham DE: Genome-wide RNAi screen identifies Letm1 as a mitochondrial Ca2+/H+ antiporter. Science. 2009, 326 (5949): 144-7. 10.1126/science.1175145.
Nowikovsky K, Froschauer EM, Zsurka G, Samaj J, Reipert S, Kolisek M, Wiesenberger G, Schweyen RJ: The LETM1/YOL027 gene family encodes a factor of the mitochondrial K+ homeostasis with a potential role in the Wolf-Hirschhorn syndrome. J Biol Chem. 2004, 279 (29): 30307-15. 10.1074/jbc.M403607200.
Palty R, Silverman WF, Hershfinkel M, Caporale T, Sensi SL, Parnis J, Nolte C, Fishman D, Shoshan-Barmatz V, Herrmann S, Khananshvili D, Sekler I: NCLX is an essential component of mitochondrial Na+/Ca2+ exchange. Proc Natl Acad Sci USA. 2010, 107 (1): 436-41. 10.1073/pnas.0908099107.
Brini M, Carafoli E: Calcium pumps in health and disease. Physiol Rev. 2009, 89 (4): 1341-78. 10.1152/physrev.00032.2008.
Thomas RC: The plasma membrane calcium ATPase (PMCA) of neurones is electroneutral and exchanges 2 H+ for each Ca2+ or Ba2+ ion extruded. J Physiol. 2009, 587 (Pt 2): 315-27.
Philipson KD, Nicoll DA: Sodium-calcium exchange: a molecular perspective. Annu Rev Physiol. 2000, 62: 111-33. 10.1146/annurev.physiol.62.1.111.
Van Baelen K, Vanoevelen J, Missiaen L, Raeymaekers L, Wuytack F: The Golgi PMR1 P-type ATPase of Caenorhabditis elegans. Identification of the gene and demonstration of calcium and manganese transport. J Biol Chem. 2001, 276 (14): 10683-91. 10.1074/jbc.M010553200.
Dode L, Andersen JP, Raeymaekers L, Missiaen L, Vilsen B, Wuytack F: Functional comparison between secretory pathway Ca2+/Mn2+-ATPase (SPCA) 1 and sarcoplasmic reticulum Ca2+-ATPase (SERCA) 1 isoforms by steady-state and transient kinetic analyses. J Biol Chem. 2005, 280 (47): 39124-34. 10.1074/jbc.M506181200.
Yu X, Carroll S, Rigaud JL, Inesi G: H+ countertransport and electrogenicity of the sarcoplasmic reticulum Ca2+ pump in reconstituted proteoliposomes. Biophys J. 1993, 64 (4): 1232-42. 10.1016/S0006-3495(93)81489-9.
Toyoshima C: Structural aspects of ion pumping by Ca2+-ATPase of sarcoplasmic reticulum. Arch Biochem Biophys. 2008, 476 (1): 3-11. 10.1016/j.abb.2008.04.017.
De Giorgi F, Lartigue L, Ichas F: Electrical coupling and plasticity of the mitochondrial network. Cell Calcium. 2000, 28 (5-6): 365-70. 10.1054/ceca.2000.0177.
Lombardi A, Damon M, Vincent A, Goglia F, Herpin P: Characterisation of oxidative phosphorylation in skeletal muscle mitochondria subpopulations in pig: a study using top-down elasticity analysis. FEBS Lett. 2000, 475 (2): 84-8. 10.1016/S0014-5793(00)01633-1.
Collins TJ, Berridge MJ, Lipp P, Bootman MD: Mitochondria are morphologically and functionally heterogeneous within cells. EMBO J. 2002, 21 (7): 1616-27. 10.1093/emboj/21.7.1616.
Gomes LC, Di Benedetto G, Scorrano L: During autophagy mitochondria elongate, are spared from degradation and sustain cell viability. Nat Cell Biol. 2011, 13 (5): 589-98. 10.1038/ncb2220.
Pinton P, Giorgi C, Siviero R, Zecchini E, Rizzuto R: Calcium and apoptosis: ER-mitochondria Ca2+ transfer in the control of apoptosis. Oncogene. 2008, 27 (50): 6407-18. 10.1038/onc.2008.308.
Rimessi A, Giorgi C, Pinton P, Rizzuto R: The versatility of mitochondrial calcium signals: from stimulation of cell metabolism to induction of cell death. Biochim Biophys Acta. 2008, 1777 (7-8): 808-16. 10.1016/j.bbabio.2008.05.449.
Rizzuto R, Brini M, Murgia M, Pozzan T: Microdomains with high Ca2+ close to IP3-sensitive channels that are sensed by neighboring mitochondria. Science. 1993, 262 (5134): 744-7. 10.1126/science.8235595.
Szabadkai G, Bianchi K, Várnai P, De Stefani D, Wieckowski MR, Cavagna D, Nagy AI, Balla T, Rizzuto R: Chaperone-mediated coupling of endoplasmic reticulum and mitochondrial Ca2+ channels. J Cell Biol. 2006, 175 (6): 901-11. 10.1083/jcb.200608073.
Hayashi T, Su TP: Sigma-1 receptor chaperones at the ER-mitochondrion interface regulate Ca2+ signaling and cell survival. Cell. 2007, 131 (3): 596-610. 10.1016/j.cell.2007.08.036.
Marchi S, Rimessi A, Giorgi C, Baldini C, Ferroni L, Rizzuto R, Pinton P: Akt kinase reducing endoplasmic reticulum Ca2+ release protects cells from Ca2+-dependent apoptotic stimuli. Biochem Biophys Res Commun. 2008, 375 (4): 501-5. 10.1016/j.bbrc.2008.07.153.
Boehning D, Patterson RL, Sedaghat L, Glebova NO, Kurosaki T, Snyder SH: Cytochrome c binds to inositol (1,4,5) trisphosphate receptors, amplifying calcium-dependent apoptosis. Nat Cell Biol. 2003, 5 (12): 1051-61. 10.1038/ncb1063.
Giorgio M, Migliaccio E, Orsini F, Paolucci D, Moroni M, Contursi C, Pelliccia G, Luzi L, Minucci S, Marcaccio M, Pinton P, Rizzuto R, Bernardi P, Paolucci F, Pelicci PG: Electron transfer between cytochrome c and p66Shc generates reactive oxygen species that trigger mitochondrial apoptosis. Cell. 2005, 122 (2): 221-33. 10.1016/j.cell.2005.05.011.
Pinton P, Rimessi A, Marchi S, Orsini F, Migliaccio E, Giorgio M, Contursi C, Minucci S, Mantovani F, Wieckowski MR, Del Sal G, Pelicci PG, Rizzuto R: Protein kinase C beta and prolyl isomerase 1 regulate mitochondrial effects of the life-span determinant p66Shc. Science. 2007, 315 (5812): 659-63. 10.1126/science.1135380.
Lebiedzinska M, Karkucinska-Wieckowska A, Giorgi C, Karczmarewicz E, Pronicka E, Pinton P, Duszynski J, Pronicki M, Wieckowski MR: Oxidative stress-dependent p66Shc phosphorylation in skin fibroblasts of children with mitochondrial disorders. Biochim Biophys Acta. 2010, 1797 (6-7): 952-60. 10.1016/j.bbabio.2010.03.005.
Lebiedzinska M, Duszynski J, Rizzuto R, Pinton P, Wieckowski MR: Age-related changes in levels of p66Shc and serine 36-phosphorylated p66Shc in organs and mouse tissues. Arch Biochem Biophys. 2009, 486 (1): 73-80. 10.1016/j.abb.2009.03.007.
Giorgi C, Ito K, Lin HK, Santangelo C, Wieckowski MR, Lebiedzinska M, Bononi A, Bonora M, Duszynski J, Bernardi R, Rizzuto R, Tacchetti C, Pinton P, Pandolfi PP: PML regulates apoptosis at endoplasmic reticulum by modulating calcium release. Science. 2010, 330 (6008): 1247-51. 10.1126/science.1189157.
Wieckowski MR, Szabadkai G, Wasilewski M, Pinton P, Duszyński J, Rizzuto R: Overexpression of adenine nucleotide translocase reduces Ca2+ signal transmission between the ER and mitochondria. Biochem Biophys Res Commun. 2006, 348 (2): 393-9. 10.1016/j.bbrc.2006.07.072.
Breckenridge DG, Stojanovic M, Marcellus RC, Shore GC: Caspase cleavage product of BAP31 induces mitochondrial fission through endoplasmic reticulum calcium signals, enhancing cytochrome c release to the cytosol. J Cell Biol. 2003, 160 (7): 1115-27. 10.1083/jcb.200212059.
John LM, Lechleiter JD, Camacho P: Differential modulation of SERCA2 isoforms by calreticulin. J Cell Biol. 1998, 142 (4): 963-73. 10.1083/jcb.142.4.963.
Higo T, Hattori M, Nakamura T, Natsume T, Michikawa T, Mikoshiba K: Subtype-specific and ER lumenal environment-dependent regulation of inositol 1,4,5-trisphosphate receptor type 1 by ERp44. Cell. 2005, 120 (1): 85-98. 10.1016/j.cell.2004.11.048.
Gilady SY, Bui M, Lynes EM, Benson MD, Watts R, Vance JE, Simmen T: Ero1alpha requires oxidizing and normoxic conditions to localize to the mitochondria-associated membrane (MAM). Cell Stress Chaperones. 2010, 15 (5): 619-29. 10.1007/s12192-010-0174-1.
Rapizzi E, Pinton P, Szabadkai G, Wieckowski MR, Vandecasteele G, Baird G, Tuft RA, Fogarty KE, Rizzuto R: Recombinant expression of the voltage-dependent anion channel enhances the transfer of Ca2+ microdomains to mitochondria. J Cell Biol. 2002, 159 (4): 613-24. 10.1083/jcb.200205091.
Simmen T, Aslan JE, Blagoveshchenskaya AD, Thomas L, Wan L, Xiang Y, Feliciangeli SF, Hung CH, Crump CM, Thomas G: PACS-2 controls endoplasmic reticulum-mitochondria communication and Bid-mediated apoptosis. EMBO J. 2005, 24 (4): 717-29. 10.1038/sj.emboj.7600559.
Cui Z, Vance JE, Chen MH, Voelker DR, Vance DE: Cloning and expression of a novel phosphatidylethanolamine N-methyltransferase. A specific biochemical and cytological marker for a unique membrane fraction in rat liver. J Biol Chem. 1993, 268 (22): 16655-63.
Rusiñol AE, Cui Z, Chen MH, Vance JE: A unique mitochondria-associated membrane fraction from rat liver has a high capacity for lipid synthesis and contains pre-Golgi secretory proteins including nascent lipoproteins. J Biol Chem. 1994, 269 (44): 27494-502.
Roderick HL, Lechleiter JD, Camacho P: Cytosolic phosphorylation of calnexin controls intracellular Ca2+ oscillations via an interaction with SERCA2b. J Cell Biol. 2000, 149 (6): 1235-48. 10.1083/jcb.149.6.1235.
Chami M, Oulès B, Szabadkai G, Tacine R, Rizzuto R, Paterlini-Bréchot P: Role of SERCA1 truncated isoform in the proapoptotic calcium transfer from ER to mitochondria during ER stress. Mol Cell. 2008, 32 (5): 641-51. 10.1016/j.molcel.2008.11.014.
Ardail D, Popa I, Bodennec J, Louisot P, Schmitt D, Portoukalian J: The mitochondria-associated endoplasmic-reticulum subcompartment (MAM fraction) of rat liver contains highly active sphingolipid-specific glycosyltransferases. Biochem J. 2003, 371 (Pt 3): 1013-9.
Kuge O, Yamakawa Y, Nishijima M: Enhancement of transport-dependent decarboxylation of phosphatidylserine by S100B protein in permeabilized Chinese hamster ovary cells. J Biol Chem. 2001, 276 (26): 23700-6. 10.1074/jbc.M101911200.
Voelker DR: Bridging gaps in phospholipid transport. Trends Biochem Sci. 2005, 30 (7): 396-404. 10.1016/j.tibs.2005.05.008.
Schmitt M, Grand-Perret T: Regulated turnover of a cell surface-associated pool of newly synthesized apolipoprotein E in HepG2 cells. J Lipid Res. 1999, 40 (1): 39-49.
Hayashi T, Su TP: Regulating ankyrin dynamics: Roles of sigma-1 receptors. Proc Natl Acad Sci USA. 2001, 98 (2): 491-6. 10.1073/pnas.021413698.
Myhill N, Lynes EM, Nanji JA, Blagoveshchenskaya AD, Fei H, Carmine-Simmen K, Cooper TJ, Thomas G, Simmen T: The subcellular distribution of calnexin is mediated by PACS-2. Mol Biol Cell. 2008, 19 (7): 2777-88. 10.1091/mbc.E07-10-0995.
Pinton P, Rizzuto R: Bcl-2 and Ca2+ homeostasis in the endoplasmic reticulum. Cell Death Differ. 2006, 13 (8): 1409-18. 10.1038/sj.cdd.4401960.
Ardail D, Gasnier F, Lermé F, Simonot C, Louisot P, Gateau-Roesch O: Involvement of mitochondrial contact sites in the subcellular compartmentalization of phospholipid biosynthetic enzymes. J Biol Chem. 1993, 268 (34): 25985-92.
Eilers M, Endo T, Schatz G: Adriamycin, a drug interacting with acidic phospholipids, blocks import of precursor proteins by isolated yeast mitochondria. J Biol Chem. 1989, 264 (5): 2945-50.
Bozidis P, Williamson CD, Colberg-Poley AM: Mitochondrial and secretory human cytomegalovirus UL37 proteins traffic into mitochondrion-associated membranes of human cells. J Virol. 2008, 82 (6): 2715-26. 10.1128/JVI.02456-07.
Sheikh MY, Choi J, Qadri I, Friedman JE, Sanyal AJ: Hepatitis C virus infection: molecular pathways to metabolic syndrome. Hepatology. 2008, 47 (6): 2127-33. 10.1002/hep.22269.
Li G, Mongillo M, Chin KT, Harding H, Ron D, Marks AR, Tabas I: Role of ERO1-alpha-mediated stimulation of inositol 1,4,5-triphosphate receptor activity in endoplasmic reticulum stress-induced apoptosis. J Cell Biol. 2009, 186 (6): 783-92. 10.1083/jcb.200904060.
Acknowledgements
This research was supported by: the Ministry of Science and Higher Education, Poland, grants N301 092 32/3407, N407 075 137, by the Polish Mitochondrial Network for MRW, JD and JS.
And by the Italian Association for Cancer Research (AIRC), Telethon (GGP09128), local funds from the University of Ferrara, the Italian Ministry of Education, University and Research (COFIN), the Italian Cystic Fibrosis Research Foundation and Italian Ministry of Health to P.P.
SM was supported by a FIRC fellowship; AB was supported by a research fellowship FISM - Fondazione Italiana Sclerosi Multipla - Cod. 2010/B/1; SP was supported by a training fellowship FISM - Fondazione Italiana Sclerosi Multipla - Cod. 2010/B/13; JS was supported by PhD fellowship from The Foundation for Polish Science (FNP), UE, European Regional Development Fund and Operational Programme "Innovative economy".
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
SP, JMS, CA, AB, MB, EDM, CG, SM, SM, FP, AR, JD, MRW and PP contributed to the preparation of the manuscript and approval of its final version.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Patergnani, S., Suski, J.M., Agnoletto, C. et al. Calcium signaling around Mitochondria Associated Membranes (MAMs). Cell Commun Signal 9, 19 (2011). https://doi.org/10.1186/1478-811X-9-19
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1478-811X-9-19