
Abstract
Norovirus (NoV) was first reported as causative agent of gastroenteritis in 1972, when students and staff of an elementary school located in Norwalk, Ohio, USA, presented vomitus and diarrhoea. After this, a number of studies definitively associated the NoV infection with outbreaks of acute gastroenteritis. Nowadays, NoV is considered the leading cause of nonbacterial gastroenteritis outbreaks and severe childhood diarrhoea worldwide, including water- and food-borne outbreaks. In animals, NoV infections have been reported in swine, bovine, ovine, canine, feline, and murine. The pathogenic role of NoV infection and its impact in animal health are not completely clear. Most of the epidemiological studies detected NoV in asymptomatic animal hosts worldwide. However, there are studies that associated the NoV infection with disease, especially enteritis, in swine, bovine, canine, and feline. This chapter presents the NoV properties and describes the pathogenesis, epidemiology, and available techniques for diagnosis of the virus infection. Additionally, the NoV infection management and prophylaxis measures for livestock animal species are approached.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
Norovirus (NoV) was first reported as causative agent of gastroenteritis in 1972, when students and staff of an elementary school located in Norwalk, Ohio, USA, presented vomitus and diarrhoea (Atmar and Estes 2001). After this, a number of studies definitively associated the NoV infection with outbreaks of acute gastroenteritis (Karst et al. 2015). Nowadays, NoV is considered the leading cause of nonbacterial gastroenteritis outbreaks and severe childhood diarrhoea worldwide, including water- and food-borne outbreaks (Patel et al. 2009; Karst et al. 2014).
In animals NoV infections have been reported in swine, bovine, ovine, canine, feline, and murine. The pathogenic role of NoV infection and its impact in animal health are not completely clear. Most of the epidemiological studies detected NoV in asymptomatic animal hosts worldwide (van der Poel et al. 2000; Keum et al. 2009; L’Homme et al. 2009). However, there are studies that associated the NoV infection with disease, especially enteritis, in canine (Martella et al. 2008), feline (Martella et al. 2007), swine (Shen et al. 2012), and bovine (Otto et al. 2011).
1 The Norovirus
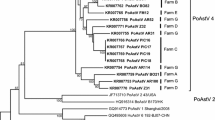
The Caliciviridae family comprises the genera Norovirus, Lagovirus, Nebovirus, Sapovirus, and Vesivirus (Fig. 6.1). Norwalk virus, previously named Norovirus, is the single representative species of Norovirus genus (ICTV 2014).

Genetic relationship of virus members of Caliciviridae family, including the representative strains of each of the five genera. The GenBank accession numbers of the sequences are indicated between parentheses. BEC bovine enteric calicivirus, RHVCG rabbit haemorrhagic disease virus – calicivirus genome, EBHSV European brown hare syndrome virus, PEC porcine enteric calicivirus, FCV feline calicivirus, VESV vesicular exanthema of swine virus
Members of Norovirus genus are small, non-enveloped, with an icosahedral capsid, and present 27–40 nm in diameter (Fig. 6.2). Virus particle’s buoyant density is 1.33–1.41 g/cm3 in caesium chloride gradient (Kapikian et al. 1973). The NoV has a linear, single-stranded, positive-sense RNA genome of approximately 7.5 kb in size. The 3′ end of the RNA genome is polyadenylated, and the 5′ end has a covalently linked protein, named VPg. This protein acts primarily in the replication process by binding initiation factors for the translation of the virus RNA.

Electron microscopy of norovirus particles present in human faecal samples stained with 2 % potassium phosphotungstate (Courtesy of Dr. J. Vinje, National Calicivirus Laboratory, Centers of Disease Control and Prevention (CDC), Atlanta, GA, USA)
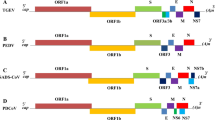
The genome of NoV is organised into three open reading frames (ORFs). The ORF1 encodes a polyprotein with approximately 200 kDa that is cleaved by the cysteine proteinase, encoded by the virus genome. The cleaved polyprotein gives origin to six nonstructural proteins, including the 2C helicase, 3C protease, and 3D RNA-dependent RNA polymerase (RdRp). The ORF2 and ORF3 encode two structural proteins that are, respectively, the VP1, which is the major capsid protein, and the VP2, a minor structural protein that is responsible for the virus genome packaging. The VP1 is functionally divided into shell (S) and protrusion (P) domains. The last is divided into P1, which is formed by two noncontiguous regions, and the hypervariable P2 that is inserted between the P1 subregions (Fig. 6.3) (Jiang et al. 1993; Ettayebi and Hardy 2003). A fourth ORF, tentatively named virulence factor 1 (VF1), which encodes a protein involved in the regulation of innate immune response has been described in the murine NoV genome (McFadden et al. 2011).

Schematic genome organization of Norovirus. Open reading frames (ORFs) and the respective virus proteins for each genomic region are shown. ORF1 encodes a polyprotein that is cleaved into nonstructural proteins. ORF2 and ORF3 encode the structural proteins VP1 and VP2, respectively, both composing the virus capsid. The VP1 is divided into shell (S) and protrusion (P) domains. P domain is externally exposed
Based on the complete deduced amino acid (aa) sequence of the VP1 capsid protein, NoVs are classified into six genogroups (GI-GVI) with the proposed seventh genogroup (GVII) (Vinje 2015). Genogroups have been further subdivided into at least 31 genotypes (Fig. 6.4). Human NoV strains are organised into GI, GII, and GIV. The human NoV GII is the most commonly detected and is classified into other distinct genotypes, of which the GII.4 is the most frequently detected in the majority of the recent large outbreaks (Karst et al. 2015).

Norovirus (NoV) is classified into seven genogroups and their genotypes based on the VP1 capsid protein. GI.1 is the NoV prototype strain. *The GII.11, GII.18, and GII.19 NoV strains infect swine; GIII.1 and GIII.2 bovine; GIII.3 ovine; GIV.2 canine and feline; GV.1 mice; and GV.2 rats. The GVI and GVII NoV strains were reported in canine
Differently from the human NoV, animal NoVs are less genetically variable. The canine NoV is classified as GIV and GVI, while the feline NoV is classified as GIV (Martella et al. 2008; Pinto et al. 2012). The murine NoV strains are in the GV (Zheng et al. 2006; McFadden et al. 2011).
The bovine and ovine NoV isolates are in the genogroup GIII. The molecular characterisation of bovine NoV genome showed that there are two different bovine NoV genotypes within the GIII genogroup. The prototypes were named Jena agent (Bo/Jena/80/DE) for the genotype 1 and Newbury agent-2 (Bo/Newbury2/76/UK) for the genotype 2 (Di Martino et al. 2014).
Porcine NoV strains are classified into GII and are closest to the most prevalent human NoV isolates. The porcine NoV GII strains detected to date are distributed into the three genotypes GII.11, .18, and .19, while the human NoV GII is classified into other distinct genotypes (Zheng et al. 2006). Since the porcine NoV strains have grouped only into these three genotypes, it was suggested that these strains have been adapted and its occurrence is restricted to pigs (L’Homme et al. 2009). However, the porcine NoV GII.18 was shown to be genetically and antigenically most closely related to human GII NoV, raising questions of whether pigs may be reservoirs for emergence of new human NoV strains (Wang et al. 2005). Figure 6.5 presents a dendrogram showing the NoV classification into genotypes and their genetic relationship.

Norovirus (NoV) classification into genotypes based on the VP1 capsid protein (From Vinje (2015), Copyright© American Society for Microbiology (2015), with permission)
2 Pathogenesis of the Disease
NoV infection can be symptomatic or asymptomatic; however, the pathogenesis of norovirus in humans and animals is not fully clarified. The incubation period after exposure to NoV is short, varying between 24 and 48 h. Symptoms of infection include acute enteritis with non-haemorrhagic diarrhoea, vomitus (characteristic sign in outbreaks), nausea, anorexia, abdominal pain, and mild fever. However, asymptomatic infections occur in one third of individuals experimentally infected. The disease duration is short (12–60 h) and self-limiting; however, immunosuppressed patients may have chronic diarrhoea and excrete viruses for months or years (Karst et al. 2015).
Previous studies with NoV performed in volunteers showed that the target cells for virus replication are primarily enterocytes of the proximal portion of the intestine, leading to malabsorption diarrhoea. Although the intestinal epithelium appears to remain intact, there are specific histopathological lesions by infection of human NoV in the jejunum, including atrophy in intestinal villi, breakdown of intestinal epithelial cells, hyperplasia crypt cells, and vacuolated and mononuclear inflammatory infiltrate in the lamina propria of villi. The malabsorption is related to the shortened microvilli and decreased brush border enzyme activity, both observed in acute infection (Karst et al. 2015).
Bovine NoV has been detected in diarrhoeic and non-diarrhoeic cattle faecal samples. Newborn calves that were inoculated with the bovine NoV GIII.1 (Jena agent) by the oral route presented severe diarrhoea in a very short incubation period (Otto et al. 2011). Experimental infection with the bovine NoV GIII.2 (Newbury agent-2) showed calves presenting diarrhoea 3–4 days postinoculation (dpi), with short duration (1 day), and no diarrhoea was observed after the re-challenge of the calves (Jor et al. 2010). This and other bovine NoV experimental-based studies showed that infected calves presented reduced appetite at the fourth and fifth dpi, non-haemorrhagic enteritis, mild to moderate diarrhoea, transient anorexia, and/or xylose malabsorption; discrete or no diarrhoea was observed in conventionally kept calves at 1–8 days of age. The rectal temperatures were between 37 and 40 °C, with pulse and respiratory rates kept within normal ranges (Jor et al. 2010; Otto et al. 2011). Infections with both bovine NoV genotypes lead to the villus atrophy and crypt hyperplasia in the proximal small intestine (Hall et al. 1984; Otto et al. 2011).
Other enteric virus agents, such as bovine rotavirus and Torovirus, infect primarily the tips and bases of villi; however, an experimental study with the bovine NoV GIII.1 showed that this virus infects all the enteroabsorptive cells. Since the bovine NoV GIII.1 causes severe villus atrophy and loss of mature enterocytes, it was suggested that these facts may limit the infection duration due to the reduction in the number of susceptible cells to the virus infection (Otto et al. 2011). On the other hand, the bovine NoV GIII.2 experimental studies showed that calves shed the NoV in faeces for at least 30 days after inoculation, regardless of the faecal consistency (diarrhoeic or not) and the duration of the clinical signs (Jor et al. 2010; Jung et al. 2014). Diarrhoea and prolonged faecal shedding of bovine NoV GIII.2 were observed even in calves that were not presented with major histological changes in the intestine, including no necrosis of intestinal epithelium, villous atrophy, or inflammatory lesions (Jung et al. 2014).
The importance of the porcine NoV as diarrhoea-causative agent in pigs is not yet fully understood. In an experimental challenge of piglets, the porcine NoV incubation period was of only 1 dpi, and the diarrhoea persisted for 2–6 days. Piglets presented mild to moderate villous atrophy and mild to moderate and multifocal villous fusion in the small intestine (Shen et al. 2012).
Previously, an experimental study inoculated piglets with the human NoV strain GII.4. The incubation period varied from 24 to 48 h; the diarrhoea was mild and self-limiting, persisting for 1–3 days. As well, the virus shedding was shown to be short, from 1 to 4 days. The virus antigen was detected in the cytoplasm of the small intestine cells. The histopathological lesions that were multifocal atrophy of the intestinal villi, enterocytes infected with low columnar morphology, and oedema of the lamina propria duodenal occurred at low frequency and were considered to be of low intensity. Another finding of this study was the increase in the number of apoptotic enterocytes (Cheetham et al. 2006).
Replication of NoV may not be restricted to enterocytes. Of all the potential experimental models studied to better understand the pathogenesis of noroviruses, the only norovirus which replicates in vitro is the murine NoV. This agent replicates in macrophages and dendritic cells derived from cultures of bone marrow cells and in mouse macrophage cell lines (RAW 264.7) (Wobus et al. 2004). The murine NoV-1 infection in knockout mice for recombination-activating gene 2 (RAG2) and signal transducer and activator of transcription 1 (STAT-1) genes, RAG2/STAT-1, showed tropism for haematopoietic cell (macrophages and dendritic cells) and development of systemic disease. Clinical signs include pneumonia, hepatitis, encephalitis, and vasculitis in brain capillaries and can be observed even in inoculation in serial passages (Karst et al. 2003; Wobus et al. 2006; Scipioni et al. 2008b). It was also demonstrated that the murine NoV can naturally infect wild and immunodeficient mice. The infection also occurs following oral or intranasal inoculation. However, although other strains of murine NoV have already been isolated from faecal samples of infected mice, it is not yet clear whether this virus is an effective enteric pathogen in this animal species (Hsu et al. 2005).
3 Epidemiology
The transmission of NoV occurs predominantly by the faecal-oral route for both human and animals. Differently of other viruses that depend on high virus concentrations for causing disease, NoV requires a low infectious dose (<10–100 virions) to establish the infection (Atmar and Estes 2006).
NoV showed a long-term survival in suspensions at environmental temperature, indicating that transmission by routes involving surface or drinking water, moisture fomites, or workplace surfaces is possible (Duizer et al. 2004). Additionally, the respiratory tract has been considered as another natural route of NoV transmission by the inhalation of aerosolised particles in vomitus (Atmar and Estes 2006).
NoV is considered a waterborne virus of primary concern, together with other virus agents, such as hepatitis A virus, hepatitis E virus, adenovirus, astrovirus, enterovirus, and rotavirus (Gibson 2014). Outbreaks of human NoV infection were associated with contaminated drinking water in different countries (Duizer et al. 2004). Studies revealed a widespread occurrence of human enteric viruses in both individual and municipal wells, showing that groundwater can be pathogen contaminated, including with NoV, and that groundwater-sourced public water systems producing water without disinfection can represent a risk of waterborne illness (Gibson 2014), including for animals.
The faecal contamination in water, food, and fomites and the direct individual-to-individual contacts are responsible for the major occurrence of gastroenteritis outbreaks determined by NoV. Since the surface/drinking water and groundwater quality can be affected by multiple sources of pathogens, vegetables also can be contaminated with NoV by irrigation with contaminated water. The food may be contaminated with virus particles since its production or crop, as in the case of oysters and fresh produce, or is contaminated on site preparation by means of handling by infected people, as in the case of cold food, sandwiches, and salads (Atmar and Estes 2006).
High rates of secondary attacks (≥30 %) among people who had contact with infected individuals lead the outbreak amplification in places where there is overcrowding, such as hospital wards, cruise ships, and shelters (Atmar and Estes 2006). In addition to the low infectious dose required for NoV transmission, continuous NoV infection is a result of the difficult elimination of the virus due to its resistance to disinfectants and many chemical products, the facilitated survival by organic debris of the clinical specimens (faeces/vomitus) in which the virus is shed, and the NoV aggregate formation that protects the virus from the environmental conditions (Kotwal and Cannon 2014).
In symptomatic animals, the virus shedding appears shortly before or during the first clinical signs and is prolonged, even after resolving of the symptoms (Scipioni et al. 2008b); the individuals with asymptomatic NoV infection also shed the virus. The period of virus shedding may range between 5 and 60 days, with a medium of 30 days. In human NoV infection, the virus is excreted in high amounts; the peak of virus RNA titres may vary from 109 to 1012 genomic copies per gramme stool and may be 1–2 log lower in symptomatic and asymptomatic individuals, respectively (Newman and Leon 2015). These facts and the NoV infectious stability for weeks or months in the environment may facilitate the NoV persistence and the virus transmission among infected and susceptible hosts (Mathijs et al. 2012).
The murine NoV is one of the most prevalent pathogens of murine, being a causative agent of systemic infection and lethal disease in immunodeficient laboratory mice (Karst et al. 2003). However, murine NoV strains were also identified from immunocompetent laboratory mice with silent infection (Hsu et al. 2005; Wobus et al. 2006). As the murine NoV is the only norovirus that replicates in cell culture, this virus is considered an excellent model to comprehend the basic mechanisms of norovirus replication in vitro and in vivo (Wobus et al. 2006).
Dogs were first evidenced to be susceptible to NoV infection in 2007 (Martella et al. 2008). Later, serological and molecular studies showed that canine NoV is spread in dogs from Europe (Ntafis et al. 2010; Caddy et al. 2013; Mesquita et al. 2014) and Asia (Tse et al. 2012; Soma et al. 2015). Norovirus also was detected from enteric disease-affected feline of Italy (Martella et al. 2007), Japan (Soma et al. 2015; Takano et al. 2015), the United States (USA (Pinto et al. 2012), and Brazil (Castro et al. 2015). Although canine and feline noroviruses are likely worldwide disseminated, their pathogenic importance has to be further studied.
Epidemiological studies have demonstrated that noroviruses are spread in livestock animals worldwide. Enteric caliciviruses morphologically similar to the human NoV were identified from diarrhoeic calves in the United Kingdom and Germany in 1978 and 1980, respectively (Woode and Bridger 1978; Gunther and Otto 1987), and were molecularly classified as bovine NoV in 1999 (Liu et al. 1999). Since then, bovine NoV has been detected in diarrhoeic and non-diarrhoeic faecal samples of beef and dairy cattle with young animals being more frequently described with the infection (Table 6.1). High seroprevalence of bovine NoV has been reported from cattle herds in Europe and North America (Table 6.2). The bovine NoV GIII.2 was prevalent in most of the studies.
The porcine NoV was first reported in Japan, where the virus RNA was recovered from caecum content of asymptomatic pigs (Sugieda et al. 1998). Subsequently, porcine NoV was reported from faecal samples of diarrhoeic and non-diarrhoeic pigs at different ages in European, Asian, Oceania, and American countries. Table 6.3 shows the frequency of porcine NoV detection by molecular assays, primarily the reverse transcription-polymerase chain reaction (RT-PCR), in different countries, and the age group studied, the number of samples evaluated, the number of positive animals, and the prevalence found. Older pigs have been more frequently detected with NoV infection; however, the virus also has been reported from suckling piglets and nursing pigs. A seroprevalence-based study showed that NoV infection is common in domestic pigs, with 71 % (78/110) and 36 % (95/226) of pig serum samples from the United States and Japan, respectively, presenting antibodies against porcine NoV (Farkas et al. 2005).
In both bovine and swine species, NoV infections with other enteric virus agents may occur. Co-infections with rotavirus, coronavirus, bovine viral diarrhoea virus (Park et al. 2007), circovirus (Shen et al. 2012), torque teno sus virus (Leme et al. 2014, unpublished data), and even with other caliciviruses (Hassine-Zaafrane et al. 2012) were reported.
To date, there are limited epidemiological data regarding NoV infection in ovine. In 2007, a study conducted in New Zealand screened sheep for the virus. In that case, animals that were positive for NoV were not presenting clinical signs suggestive of the infection. Another study was conducted in Belgium in the same year; however, sheep were not detected with the virus (Mathijs et al. 2012).
Interestingly, a seasonality pattern of NoV infection has been reported. Although the infection can occur in different seasons, wintertime seasonality is observed for different host species, including livestock animals (Hassine-Zaafrane et al. 2012; Ahmed et al. 2013; Silva et al. 2015).
3.1 Zoonotic Transmission
A concern regarding NoV infection is the possibility of cross infection among human and animal NoV strains, which raises questions about the zoonotic transmission potential of noroviruses. Studies based on the molecular epidemiology of noroviruses have shown a closer genetic relationship of human and animal noroviruses and the emergence of recombinant NoV strains in different hosts (Koopmans 2008).
Serological surveys have revealed antibodies against animal NoV in humans, including general population and veterinarians (Widdowson et al. 2005; Menon et al. 2013), and antibodies against human NoV in swine (Farkas et al. 2005). Molecular studies also revealed the presence of human NoV strains, including the most prevalent in the major recent large epidemics (GII.4), in cattle, in pig faeces, and in raw pork meat (Mattison et al. 2007). Additionally, the experimental challenge of gnotobiotic calves and piglets with human NoV showed virus replication and seroconversion (Cheetham et al. 2006; Souza et al. 2008).
Although the zoonotic transmission is likely, this hypothesis has not been proved. A possible explanation for the detection of antibodies against animal NoVs in human may be the existence of cross-reactive epitopes between the different NoV strains, as shown between human and bovine NoVs (Scipioni et al. 2008b). Noroviruses are thought to be a species-specific pathogen (Karst et al. 2015), and further studies are needed to fully comprehend the role of animals as reservoirs for human NoV infection.
4 Progress in Diagnosis and Management of NoV Infection
4.1 Diagnostic Tests
The techniques used for the laboratory diagnosis of both human and animal NoVs include the direct detection of intact virus particles by electron microscopy (EM) and of the virus RNA by RT-PCR. Currently, the development of other techniques for detecting and quantifying the NoV RNA by RT-quantitative PCR (RT-qPCR) has been reported for the diagnosis of human (Vinje 2015), swine (Machnowska et al. 2014), and bovine (Jor et al. 2010; Yilmaz et al. 2011) NoV strains. Additionally, the next-generation sequencing (NGS) has been a useful tool for the identification of evolutionary changes in relation to NoV epidemiological data (Bavelaar et al. 2015). Immunoassays, such as enzyme-linked immunosorbent assay (ELISA), are able to detect viral antigens or antibodies, both techniques being associated with the production of recombinant virus-like particles (VLPs) (Wang et al. 2007; Mauroy et al. 2009). Other techniques, such as microarray hybridisation (Wang et al. 2006; Scheuer et al. 2013) and immunohistochemistry (Otto et al. 2011), may also be used for the virus diagnosis. Table 6.4 presents the advantages and disadvantages of techniques used for the laboratory diagnosis of NoV infection.
Samples
In general, faecal samples of clinically affected animals present higher concentrations of NoV particles, and, therefore, faeces are considered the best samples for diagnosis of the infection, although the virus can be detected from rectal swabs and vomitus (Vinje 2015). The first reports of NoV infections were based on EM, since other diagnostic methods were not yet available for the virus detection at that time. However, NoV particles are morphologically similar to other small round viruses, such as sapovirus, rotavirus, astrovirus, and enteric picornavirus, and the differentiation among the viruses is harder (Wang et al. 2007). The immune EM (IEM) and solid-phase IEM are variations of the technique that are based on antigen-antibody reaction and visualised by negative staining; both can be used for NoV detection. Nowadays, the EM is generally used for research purposes and not for routine diagnosis of NoV infection.
RT-PCR and RT-qPCR
Since the first reports of NoV from different hosts, the development of diagnostic methods has been hampered by the lack of a cell culture system for NoVs, other than murine NoV (Scipioni et al. 2008b). With the development of the molecular tools for NoV diagnosis, such as conventional or endpoint RT-PCR in the mid-1990s and later the RT-qPCR and their variations, an increased number of NoV sequences became available, and the infection diagnosis became faster and trusty (Vinje 2015).
Among the factors that can affect the sensibility and/or specificity of the RT-PCR is the nucleic acid extraction method, which has to efficiently recover the virus RNA and remove the RT-PCR inhibitors. The conserved RdRp gene is frequently targeted in the RT-PCR assays for the virus diagnosis, while the VP1 region is used for the virus genotyping. The highly variable NoV strains become the oligonucleotide primer selection a hard work, since a single primer pair likely will not be able to detect all the circulating NoV strains (Wang et al. 2007). Although several primer sets can be used for the detection of genetically diverse virus strains, the design of oligonucleotide primers based on the regional NoV strains in circulation is reasonable and acceptable. For this, molecular epidemiological surveillance-based studies should be conducted.
A second round of the RT-PCR, a semi-nested or a nested PCR, can be performed to increase the sensitivity of the reaction. The increased sensitivity with the use of a nested PCR can range from 10 to 1000 times; however, this technique also increases the risk of cross-contamination (Wang et al. 2007) and its use should be under closer attention.
The RT-qPCR has major advantages in comparison with the conventional RT-PCR assays, including higher sensibility and specificity, no need of agarose gel and sequencing analyses, and the ability to determine the amount of virus nucleic acid in the clinical sample. The one-step RT-qPCR is another variation of the technique, in which the reverse transcription and cDNA amplification are performed in a single reaction. With this, less sample handling is required, which decreases the risk of cross-contamination (Vinje 2015). However, this technique does not allow depth phylogenetic studies for NoV genotyping, and the conventional RT-PCR assay is required to complement the genomic analysis.
The RT-qPCR assays have been considered the gold standard for the rapid and sensitive detection of NoV in faeces, vomitus, and serum samples of clinically affected persons, as well as in food, water, and environmental samples (Vinje 2015). Nevertheless, these techniques are not largely used by the veterinary diagnostic laboratories. For the detection of NoV in bovine, both SYBR Green- (Park et al. 2009) and TaqMan (Scipioni et al. 2008a)-based RT-qPCR assays are available; however, the SYBR Green RT-qPCR has been more frequently used for this animal species host. For swine, although a TaqMan RT-qPCR has been validated for the NoV detection (Wolf et al. 2009), this technique has not currently being used. The most likely cause for the limited use of the RT-qPCR assays for animal NoV diagnosis is their high cost, even though the prices of equipment and reagents have decreased in the last years (Wang et al. 2007).
Since the recombination is common in NoV, the surveillance of emerging strains is important for understanding the NoV evolution and global epidemiology. For this, the NGS is a powerful tool in the detection and characterisation of different types of norovirus from clinical specimens (Bavelaar et al. 2015). However, most of the routine veterinary diagnostic laboratories are not equipped with the necessary tools or staff to perform the NGS (Bavelaar et al. 2015).
Enzyme Immunoassays (EIA)
NoV capsid proteins are expressed in baculovirus system to provide VLPs that are used as antigen in the EIA. The ELISA is the most frequently EIA used for animal NoV infection diagnosis. Hyperimmune polyclonal serum obtained after animal (mice, rabbits, guinea pigs) immunisation with VLPs is used as capture antibody for the ELISA-based detection of viral antigen (Scipioni et al. 2008b). The large amounts of viral soluble proteins in stool are likely responsible for the sensitivity of antigen ELISA similar to that of conventional RT-PCR. However, the hyperimmune anti-VLP serum is often highly specific with the homologous strains or viruses within the same genotype, which make the technique highly specific (Wang et al. 2007). This fact limits the use of the technique in diagnostic laboratories, since there are highly diverse NoV strains in circulation (Wang et al. 2007; Scipioni et al. 2008b). Despite this, antigen ELISA is useful for screening large number of samples since it is a rapid and simple test (Scipioni et al. 2008b) and can be used for epidemiological-based studies from host species in which larger antigenic and/or molecular NoV genomic variations were not identified.
Antibody ELISA
It is more broadly reactive than the antigen detection and is more suitable to identify heterotypic NoV infection (Scipioni et al. 2008b). Since this assay presents a good sensitivity and specificity for NoV-specific antibody detection, it has been largely used in human epidemiological studies. However, the cross-reaction of the baculovirus-expressed VLP antibodies against GI and GII human NoV was reported (Wang et al. 2007).
Antigen and Antibody ELISA
They are available for bovine (Deng et al. 2003; Mauroy et al. 2009) and swine (Farkas et al. 2005; Cheetham et al. 2006) NoV infection diagnosis. Nevertheless, it is important to mention that three common epitopes are shared among NoVs, of which one is in the same genogroup GI (human), the second is between GII (human, swine) and GIII (bovine), and the third between GI and GIII (Scipioni et al. 2008b). Therefore, as for human ELISA, cross-reaction among different genogroups can occur. Studies reported that VLPs of porcine NoV cross reacted with antibodies against human NoV (Farkas et al. 2005) and the presence of antibodies against bovine NoV GIII.2 in veterinarians in the Netherlands (Widdowson et al. 2005).
RT-PCR-based Multiplex Molecular Diagnosis Tests
In the recent years, commercial RT-PCR-based multiplex molecular diagnosis tests for the detection of several different multi-gastrointestinal pathogens have been developed. These diagnostic platforms are able to simultaneously detect enteric viruses, including NoV GI and GII, bacteria, and parasites. However, the interpretation of data generated by these multi-pathogen systems may be a challenge, since high number of mixed infections can be detected, making it difficult to determine which pathogen is responsible for gastroenteritis (Vinje 2015). Regardless of the advantages and disadvantages of the commercial tests, these multi-gastrointestinal diagnostic platforms are not yet available for animals.
4.2 Management of NoV Infection
Livestock animals affected with NoV infection usually present mild to moderate non-haemorrhagic diarrhoea. The NoV faecal shedding can be prolonged, even after the diarrhoea recovery and/or in symptomatic animals. In this context, closer attention should be given to the livestock and environmental managements in order to provide clinical support to the affected animals and to avoid the virus exposure and transmission to susceptible animals.
Livestock animals suspected or confirmed with NoV infection should be isolated from the other herd animals. Adequate fluid and electrolyte replacement and maintenance are key points to manage NoV-induced diarrhoea that is generally a self-limiting infection, typically resolved within days after exposure. The fluid and electrolyte therapies are easier to be implemented for calves relative to piglets. For piglets, oral products intended to re-establish the hydroelectrolytic balance can be used. In cases in which severe diarrhoea is present, the use of broad-spectrum antibiotic therapy is recommended to avoid/control secondary infections. It is likely that maternal antibodies in colostrum and milk limit infection and damage in the gut of nursing piglets. Even not preventing infection, the colostrum intake may have some impact in the bovine NoV infection by limiting the infection dissemination and reducing the severity and duration of diarrhoea (Otto et al. 2011). However, further pathogenic studies under field conditions should be performed to evaluate the effects of the colostrum on the course of the bovine and swine NoV infections.
5 Development of Vaccines and Prophylaxis Measures
There are no vaccines available against bovine and swine NoV infections. Therefore, the prophylaxis measures are fundamental to prevent the disease. Faeces removal, clean facilities, disinfection, and other sanitary precautions should control and/or prevent NoV infection and persistence in the environment. Since NoV is a waterborne pathogen, adequate water management practices should be adopted, including for the drinking water that is offered to the animals, which has to be of good quality, clean, and adequately chlorine treated. As NoVs are considered potentially zoonotic, management focused on public health also should be addressed.
In general, NoV is stable in the environment and may be resistant to inactivation with certain chemicals and heat, depending on the conditions in which both are used. NoV appears to be resistant to the effects of freeze/thaw (Nims and Plavsic 2013). Inactivation of NoV by heat is time and temperature dependent, and it should exceed 56 °C to have some reduction of NoV infectivity, with extensive inactivation requiring exposure times of 30 min or higher (Duizer et al. 2004; Nims and Plavsic 2013). More consistent and extensive inactivation is observed at temperatures in excess of 60 °C, regardless of calicivirus species or strain (Nims and Plavsic 2013).
Ultraviolet Radiation
Caliciviruses are susceptible to inactivation by ultraviolet radiation in the C range (UV-C, 254 nm) at fluence of 22–40 mJ/cm2. The UV-C radiation for the inactivation of caliciviruses appears to be less effective compared to parvovirus and circovirus that are inactivated by exposure to 8–13 mJ/cm2 and 5–11 mJ/cm2, respectively. On the other hand, the calicivirus UV-C inactivation is generally similar to those displayed by other small, non-enveloped viruses. The UV radiation in the B range (UV-B, 280–320 nm) at fluence of ~60 mJ/cm2 would be required to achieve calicivirus inactivation (Nims and Plavsic 2013).
Inactivation by Chemicals
Different chemical products can be used for the calicivirus inactivation, in compliance with specific conditions for each one (Table 6.5). The NoV particles display resistance to inactivation by low pH, with incomplete inactivation achieved at pH 2.7 for 3 h at room temperature (Duizer et al. 2004). Ethanol or hypochlorite solution contacts for a short period of time (1–10 min depending on the concentrations) also were reported to incomplete inactive caliciviruses (Duizer et al. 2004). Differences to the susceptibility to inactivation with these chemical agents may not afford adequate NoV inactivation.
Treatment of Animal Wastes
To improve the agriculture, livestock manure applications to field land are practised worldwide because of its potential as biofertilizer. Another common practice in rural areas is the artificial subsurface drainage that facilitates the crop production. However, it is an efficient means by which agricultural pollutants from field systems, including faecal pollution, can enter the surface water environment (Wilkes et al. 2014). The land application of manures should be based on safety parameters, and agricultural beneficial management practices, including waste treatment in cattle and pig farm units, should be implemented to reduce the persistence of NoV in the animal waste and to mitigate the manure-borne faecal contamination of the environment, including water resources. Although it is not clear if enteric viruses are efficiently inactivated by anaerobic biodigestion, the anaerobic biodigester system is a tool that can be used to treat livestock manure in order to provide safe biofertilizers.
References
Ahmed SM, Lopman BA, Levy K (2013) A systematic review and meta-analysis of the global seasonality of norovirus. PLoS One 8(10):e75922
Alcala AC, Hidalgo MA, Obando C, Vizzi E, Liprandi F, Ludert JE (2003) Molecular identification of bovine enteric calciviruses in Venezuela (In Spanish, with English abstract). Acta Cient Venez 54(2):148–152
Atmar RL, Estes MK (2001) Diagnosis of noncultivatable gastroenteritis viruses, the human caliciviruses. Clin Microbiol Rev 14(1):15–37
Atmar RL, Estes MK (2006) The epidemiologic and clinical importance of norovirus infection. Gastroenterol Clin N Am 35(2):275–290
Bavelaar HH, Rahamat-Langendoen J, Niesters HG, Zoll J, Melchers WJ (2015) Whole genome sequencing of fecal samples as a tool for the diagnosis and genetic characterization of norovirus. J Clin Virol 72:122–125
Caddy S, Emmott E, El-Attar L, Mitchell J, de Rougemont A, Brownlie J, Goodfellow I (2013) Serological evidence for multiple strains of canine norovirus in the UK dog population. PLoS One 8(12):e81596
Castro TX, Garcia RCNC, Fumian TM, Costa EM, Mello R, White PA, Leite JPG (2015) Detection and molecular characterization of caliciviruses (vesivirus and norovirus) in an outbreak of acute diarrhea in kittens from Brazil. Vet J 206(1):115–117
Cheetham S, Souza M, Meulia T, Grimes S, Han MG, Saif LJ (2006) Pathogenesis of a genogroup II human norovirus in gnotobiotic pigs. J Virol 80(21):10372–10381
Cunha JB, de Mendonca MC, Miagostovich MP, Leite JP (2010) First detection of porcine norovirus GII.18 in Latin America. Res Vet Sci 89(1):126–129
Deng Y, Batten CA, Liu BL, Lambden PR, Elschner M, Gunther H, Otto P, Schnurch P, Eichhorn W, Herbst W, Clarke IN (2003) Studies of epidemiology and seroprevalence of bovine noroviruses in Germany. J Clin Microbiol 41(6):2300–2305
Di Bartolo I, Ponterio E, Monini M, Ruggeri FM (2011) A pilot survey of bovine norovirus in northern Italy. Vet Rec 169(3):73
Di Bartolo I, Tofani S, Angeloni G, Ponterio E, Ostanello F, Ruggeri FM (2014) Detection and characterization of porcine caliciviruses in Italy. Arch Virol 159(9):2479–2484
Di Martino B, Di Profio F, Di Felice E, Melegari I, Ceci C, Mauroy A, Thiry E, Martella V, Marsilio F (2014) Genetic heterogeneity of bovine noroviruses in Italy. Arch Virol 159(10):2717–2722
Duizer E, Bijkerk P, Rockx B, De Groot A, Twisk F, Koopmans M (2004) Inactivation of caliciviruses. Appl Environ Microbiol 70(8):4538–4543
Ettayebi K, Hardy ME (2003) Norwalk virus nonstructural protein p48 forms a complex with the SNARE regulator VAP-A and prevents cell surface expression of vesicular stomatitis virus G protein. J Virol 77(21):11790–11797
Farkas T, Nakajima S, Sugieda M, Deng X, Zhong W, Jiang X (2005) Seroprevalence of noroviruses in swine. J Clin Microbiol 43(2):657–661
Gibson KE (2014) Viral pathogens in water: occurrence, public health impact, and available control strategies. Curr Opin Virol 4:50–57
Gunther H, Otto P (1987) Diarrhea in young calves. 7. “Zackenvirus” (Jena agent 117/80)-a new diarrhea pathogen in calves [In Germany]. Arch Exp Veterinarmed 41(6):934–938
Hall GA, Bridger JC, Brooker BE, Parsons KR, Ormerod E (1984) Lesions of gnotobiotic calves experimentally infected with a calicivirus-like (Newbury) agent. Vet Pathol 21(2):208–215
Hassine-Zaafrane M, Kaplon J, Sdiri-Loulizi K, Aouni Z, Pothier P, Aouni M, Ambert-Balay K (2012) Molecular prevalence of bovine noroviruses and neboviruses detected in central-eastern Tunisia. Arch Virol 157(8):1599–1604
Hsu CC, Wobus CE, Steffen EK, Riley LK, Livingston RS (2005) Development of a microsphere-based serologic multiplexed fluorescent immunoassay and a reverse transcriptase PCR assay to detect murine norovirus 1 infection in mice. Clin Diagn Lab Immunol 12(10):1145–1151
ICTV (2014) Virus taxonomy: 2014 release. International Committee on Taxonomy of Viruses. http://www.ictvonline.org/virustaxonomy.asp. Accessed 3 Jan 2016
Jiang X, Wang M, Wang K, Estes MK (1993) Sequence and genomic organization of Norwalk virus. Virology 195(1):51–61
Jor E, Myrmel M, Jonassen CM (2010) SYBR Green based real-time RT-PCR assay for detection and genotype prediction of bovine noroviruses and assessment of clinical significance in Norway. J Virol Methods 169(1):1–7
Jung K, Scheuer KA, Zhang Z, Wang Q, Saif LJ (2014) Pathogenesis of GIII.2 bovine norovirus, CV186-OH/00/US strain in gnotobiotic calves. Vet Microbiol 168(1):202–207
Kapikian AZ, Gerin JL, Wyatt RG, Thornhill TS, Chanock RM (1973) Density in cesium chloride of the 27 nm “8FIIa” particle associated with acute infectious nonbacterial gastroenteritis: determination by ultra-centrifugation and immune electron microscopy. Proc Soc Exp Biol Med 142(3):874–877
Karst SM, Wobus CE, Lay M, Davidson J, Virgin HW (2003) STAT1-dependent innate immunity to a Norwalk-like virus. Science 299(5612):1575–1578
Karst SM, Wobus CE, Goodfellow IG, Green KY, Virgin HW (2014) Advances in norovirus biology. Cell Host Microbe 15(6):668–680
Karst SM, Zhu S, Goodfellow IG (2015) The molecular pathology of noroviruses. J Pathol 235(2):206–216
Keum HO, Moon HJ, Park SJ, Kim HK, Rho SM, Park BK (2009) Porcine noroviruses and sapoviruses on Korean swine farms. Arch Virol 154(11):1765–1774
Koopmans M (2008) Progress in understanding norovirus epidemiology. Curr Opin Infect Dis 21(5):544–552
Kotwal G, Cannon JL (2014) Environmental persistence and transfer of enteric viruses. Curr Opin Virol 4:37–43
L’Homme Y, Sansregret R, Plante-Fortier E, Lamontagne AM, Lacroix G, Ouardani M, Deschamps J, Simard G, Simard C (2009) Genetic diversity of porcine Norovirus and Sapovirus: Canada, 2005-2007. Arch Virol 154(4):581–593
Liu BL, Lambden PR, Gunther H, Otto P, Elschner M, Clarke IN (1999) Molecular characterization of a bovine enteric calicivirus: relationship to the Norwalk-like viruses. J Virol 73(1):819–825
Machnowska P, Ellerbroek L, Johne R (2014) Detection and characterization of potentially zoonotic viruses in faeces of pigs at slaughter in Germany. Vet Microbiol 168(1):60–68
Martella V, Campolo M, Lorusso E, Cavicchio P, Camero M, Bellacicco AL, Decaro N, Elia G, Greco G, Corrente M, Desario C, Arista S, Banyai K, Koopmans M, Buonavoglia C (2007) Norovirus in captive lion cub (Panthera leo). Emerg Infect Dis 13(7):1071–1073
Martella V, Lorusso E, Decaro N, Elia G, Radogna A, D’Abramo M, Desario C, Cavalli A, Corrente M, Camero M, Germinario CA, Banyai K, Di Martino B, Marsilio F, Carmichael LE, Buonavoglia C (2008) Detection and molecular characterization of a canine norovirus. Emerg Infect Dis 14(8):1306–1308
Mathijs E, Stals A, Baert L, Botteldoorn N, Denayer S, Mauroy A, Scipioni A, Daube G, Dierick K, Herman L, Van Coillie E, Uyttendaele M, Thiry E (2012) A review of known and hypothetical transmission routes for noroviruses. Food Environ Virol 4(4):131–152
Mattison K, Shukla A, Cook A, Pollari F, Friendship R, Kelton D, Bidawid S, Farber JM (2007) Human noroviruses in swine and cattle. Emerg Infect Dis 13(8):1184–1188
Mauroy A, Scipioni A, Mathijs E, Miry C, Ziant D, Thys C, Thiry E (2008) Noroviruses and sapoviruses in pigs in Belgium. Arch Virol 153(10):1927–1931
Mauroy A, Scipioni A, Mathijs E, Saegerman C, Mast J, Bridger JC, Ziant D, Thys C, Thiry E (2009) Epidemiological study of bovine norovirus infection by RT-PCR and a VLP-based antibody ELISA. Vet Microbiol 137(3-4):243–251
McFadden N, Bailey D, Carrara G, Benson A, Chaudhry Y, Shortland A, Heeney J, Yarovinsky F, Simmonds P, Macdonald A, Goodfellow I (2011) Norovirus regulation of the innate immune response and apoptosis occurs via the product of the alternative open reading frame 4. PLoS Pathog 7(12):e1002413
Menon VK, George S, Shanti AA, Saravanabavan A, Samuel P, Ramani S, Estes MK, Kang G (2013) Exposure to human and bovine noroviruses in a birth cohort in southern India from 2002 to 2006. J Clin Microbiol 51(7):2391–2395
Mesquita JR, Delgado I, Costantini V, Heenemann K, Vahlenkamp TW, Vinje J, Nascimento MS (2014) Seroprevalence of canine norovirus in 14 European countries. Clin Vaccine Immunol 21(6):898–900
Mijovski JZ, Poljsak-Prijatelj M, Steyer A, Barlic-Maganja D, Koren S (2010) Detection and molecular characterisation of noroviruses and sapoviruses in asymptomatic swine and cattle in Slovenian farms. Infect Genet Evol 10(3):413–420
Newman KL, Leon JS (2015) Norovirus immunology: of mice and mechanisms. Eur J Immunol 45(10):2742–2757
Nims R, Plavsic M (2013) Inactivation of caliciviruses. Pharmaceuticals 3:358–392
Ntafis V, Xylouri E, Radogna A, Buonavoglia C, Martella V (2010) Outbreak of canine norovirus infection in young dogs. J Clin Microbiol 48(7):2605–2608
Oliver SL, Wood E, Asobayire E, Wathes DC, Brickell JS, Elschner M, Otto P, Lambden PR, Clarke IN, Bridger JC (2007) Serotype 1 and 2 bovine noroviruses are endemic in cattle in the United kingdom and Germany. J Clin Microbiol 45(9):3050–3052
Otto PH, Clarke IN, Lambden PR, Salim O, Reetz J, Liebler-Tenorio EM (2011) Infection of calves with bovine norovirus GIII.1 strain Jena virus: an experimental model to study the pathogenesis of norovirus infection. J Virol 85(22):12013–12021
Park SI, Jeong C, Kim HH, Park SH, Park SJ, Hyun BH, Yang DK, Kim SK, Kang MI, Cho KO (2007) Molecular epidemiology of bovine noroviruses in South Korea. Vet Microbiol 124(1-2):125–133
Park SI, Park DH, Saif LJ, Jeong YJ, Shin DJ, Chun YH, Park SJ, Kim HJ, Hosmillo M, Kwon HJ, Kang MI, Cho KO (2009) Development of SYBR Green real-time RT-PCR for rapid detection, quantitation and diagnosis of unclassified bovine enteric calicivirus. J Virol Methods 159(1):64–68
Patel MM, Hall AJ, Vinje J, Parashar UD (2009) Noroviruses: a comprehensive review. J Clin Virol 44(1):1–8
Pinto P, Wang Q, Chen N, Dubovi EJ, Daniels JB, Millward LM, Buonavoglia C, Martella V, Saif LJ (2012) Discovery and genomic characterization of noroviruses from a gastroenteritis outbreak in domestic cats in the US. PLoS One 7(2):e32739
Reuter G, Biro H, Szucs G (2007) Enteric caliciviruses in domestic pigs in Hungary. Arch Virol 152(3):611–614
Scheuer KA, Oka T, Hoet AE, Gebreyes WA, Molla BZ, Saif LJ, Wang Q (2013) Prevalence of porcine noroviruses, molecular characterization of emerging porcine sapoviruses from finisher swine in the United States, and unified classification scheme for sapoviruses. J Clin Microbiol 51(7):2344–2353
Scipioni A, Bourgot I, Mauroy A, Ziant D, Saegerman C, Daube G, Thiry E (2008a) Detection and quantification of human and bovine noroviruses by a TaqMan RT-PCR assay with a control for inhibition. Mol Cell Probes 22(4):215–222
Scipioni A, Mauroy A, Vinje J, Thiry E (2008b) Animal noroviruses. Vet J 178(1):32–45
Shen Q, Zhang W, Yang S, Chen Y, Ning H, Shan T, Liu J, Yang Z, Cui L, Zhu J, Hua X (2009) Molecular detection and prevalence of porcine caliciviruses in eastern China from 2008 to 2009. Arch Virol 154(10):1625–1630
Shen Q, Zhang W, Yang S, Yang Z, Chen Y, Cui L, Zhu J, Hua X (2012) Recombinant porcine norovirus identified from piglet with diarrhea. BMC Vet Res 8:155
Silva PF, Alfieri AF, Barry AF, de Arruda LR, Gardinali NR, van der Poel WH, Alfieri AA (2015) High frequency of porcine norovirus infection in finisher units of Brazilian pig-production systems. Trop Anim Health Prod 47(1):237–241
Soma T, Nakagomi O, Nakagomi T, Mochizuki M (2015) Detection of Norovirus and Sapovirus from diarrheic dogs and cats in Japan. Microbiol Immunol 59(3):123–128
Song YJ, Yu JN, Nam HM, Bak HR, Lee JB, Park SY, Song CS, Seo KH, Choi IS (2011) Identification of genetic diversity of porcine Norovirus and Sapovirus in Korea. Virus Genes 42(3):394–401
Souza M, Azevedo MS, Jung K, Cheetham S, Saif LJ (2008) Pathogenesis and immune responses in gnotobiotic calves after infection with the genogroup II.4-HS66 strain of human norovirus. J Virol 82(4):1777–1786
Sugieda M, Nagaoka H, Kakishima Y, Ohshita T, Nakamura S, Nakajima S (1998) Detection of Norwalk-like virus genes in the caecum contents of pigs. Arch Virol 143(6):1215–1221
Takano T, Kusuhara H, Kuroishi A, Takashina M, Doki T, Nishinaka T, Hohdatsu T (2015) Molecular characterization and pathogenicity of a genogroup GVI feline norovirus. Vet Microbiol 178(3-4):201–207
Thomas C, Jung K, Han MG, Hoet A, Scheuer K, Wang Q, Saif LJ (2014) Retrospective serosurveillance of bovine norovirus (GIII.2) and nebovirus in cattle from selected feedlots and a veal calf farm in 1999 to 2001 in the United States. Arch Virol 159(1):83–90
Tse H, Lau SK, Chan WM, Choi GK, Woo PC, Yuen KY (2012) Complete genome sequences of novel canine noroviruses in Hong Kong. J Virol 86(17):9531–9532
van der Poel WH, Vinje J, van der Heide R, Herrera MI, Vivo A, Koopmans MP (2000) Norwalk-like calicivirus genes in farm animals. Emerg Infect Dis 6(1):36–41
van der Poel WH, van der Heide R, Verschoor F, Gelderblom H, Vinje J, Koopmans MP (2003) Epidemiology of Norwalk-like virus infections in cattle in The Netherlands. Vet Microbiol 92(4):297–309
Vinje J (2015) Advances in laboratory methods for detection and typing of norovirus. J Clin Microbiol 53(2):373–381
Wang QH, Han MG, Cheetham S, Souza M, Funk JA, Saif LJ (2005) Porcine noroviruses related to human noroviruses. Emerg Infect Dis 11(12):1874–1881
Wang QH, Souza M, Funk JA, Zhang W, Saif LJ (2006) Prevalence of noroviruses and sapoviruses in swine of various ages determined by reverse transcription-PCR and microwell hybridization assays. J Clin Microbiol 44(6):2057–2062
Wang QH, Costantini V, Saif LJ (2007) Porcine enteric caliciviruses: genetic and antigenic relatedness to human caliciviruses, diagnosis and epidemiology. Vaccine 25(30):5453–5466
Widdowson MA, Rockx B, Schepp R, van der Poel WH, Vinje J, van Duynhoven YT, Koopmans MP (2005) Detection of serum antibodies to bovine norovirus in veterinarians and the general population in the Netherlands. J Med Virol 76(1):119–128
Wilkes G, Brassard J, Edge TA, Gannon V, Gottschall N, Jokinen CC, Jones TH, Khan IU, Marti R, Sunohara MD, Topp E, Lapen DR (2014) Long-term monitoring of waterborne pathogens and microbial source tracking markers in paired agricultural watersheds under controlled and conventional tile drainage management. Appl Environ Microbiol 80(12):3708–3720
Wobus CE, Karst SM, Thackray LB, Chang KO, Sosnovtsev SV, Belliot G, Krug A, Mackenzie JM, Green KY, Virgin HW (2004) Replication of Norovirus in cell culture reveals a tropism for dendritic cells and macrophages. PLoS Biol 2(12):e432
Wobus CE, Thackray LB, Virgin HW (2006) Murine norovirus: a model system to study norovirus biology and pathogenesis. J Virol 80(11):5104–5112
Wolf S, Williamson W, Hewitt J, Lin S, Rivera-Aban M, Ball A, Scholes P, Savill M, Greening GE (2009) Molecular detection of norovirus in sheep and pigs in New Zealand farms. Vet Microbiol 133(1-2):184–189
Woode GN, Bridger JC (1978) Isolation of small viruses resembling astroviruses and caliciviruses from acute enteritis of calves. J Med Microbiol 11(4):441–452
Yilmaz H, Turan N, Altan E, Bostan K, Yilmaz A, Helps CR, Cho KO (2011) First report on the phylogeny of bovine norovirus in Turkey. Arch Virol 156(1):143–147
Zheng DP, Ando T, Fankhauser RL, Beard RS, Glass RI, Monroe SS (2006) Norovirus classification and proposed strain nomenclature. Virology 346(2):312–323
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Alfieri, A.A., Leme, R.A., Alfieri, A.F. (2017). Norovirus Infection. In: Bayry, J. (eds) Emerging and Re-emerging Infectious Diseases of Livestock. Springer, Cham. https://doi.org/10.1007/978-3-319-47426-7_6
Download citation
DOI: https://doi.org/10.1007/978-3-319-47426-7_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-47424-3
Online ISBN: 978-3-319-47426-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)