Cytokine-Targeted Therapeutics for KSHV-Associated Disease


Abstract
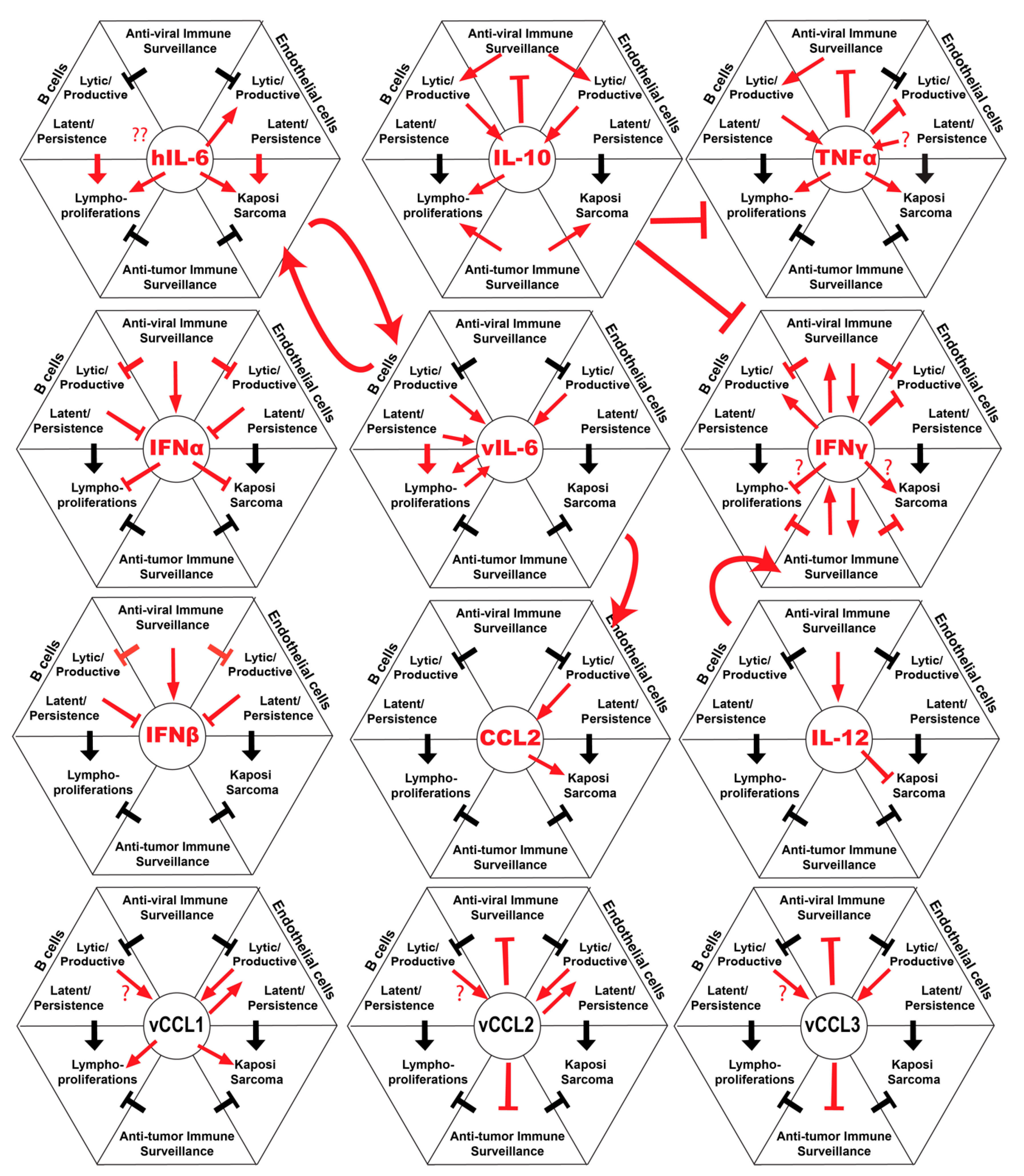
:1. Introduction
2. IL-6
3. IL-10
4. Interferons
5. TNFa
6. IL-12
7. Host and Viral Chemokines
8. Summary and Future Considerations
Author Contributions
Funding
Conflicts of Interest
References
- Chang, Y.; Cesarman, E.; Pessin, M.S.; Lee, F.; Culpepper, J.; Knowles, D.M.; Moore, P.S. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science 1994, 266, 1865–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ablashi, D.V.; Chatlynne, L.G.; Whitman, J.E.; Cesarman, E. Spectrum of Kaposi’s sarcoma-associated herpesvirus, or human herpesvirus 8, diseases. Clin. Microbiol. Rev. 2002, 15, 439–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesarman, E.; Chang, Y.; Moore, P.S.; Said, J.W.; Knowles, D.M. Kaposi’s sarcoma–associated herpesvirus-like DNA sequences in AIDS-related body-cavity–based lymphomas. N. Engl. J. Med. 1995, 332, 1186–1191. [Google Scholar] [CrossRef] [PubMed]
- Soulier, J.; Grollet, L.; Oksenhendler, E.; Cacoub, P.; Cazals-Hatem, D.; Babinet, P.; d’Agay, M.; Clauvel, J.-P.; Raphael, M.; Degos, L. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in multicentric Castleman’s disease [see comments]. Blood 1995, 86, 1276–1280. [Google Scholar] [CrossRef] [Green Version]
- Uldrick, T.S.; Wang, V.; O’Mahony, D.; Aleman, K.; Wyvill, K.M.; Marshall, V.; Steinberg, S.M.; Pittaluga, S.; Maric, I.; Whitby, D. An interleukin-6-related systemic inflammatory syndrome in patients co-infected with Kaposi sarcoma-associated herpesvirus and HIV but without Multicentric Castleman disease. Clin. Infect. Dis. 2010, 51, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Levy, J.; Ziegler, J. Acquired immunodeficiency syndrome is an opportunistic infection and Kaposi’s sarcoma results from secondary immune stimulation. Lancet 1983, 322, 78–81. [Google Scholar] [CrossRef]
- Lukac, D.M.; Renne, R.; Kirshner, J.R.; Ganem, D. Reactivation of Kaposi’s sarcoma-associated herpesvirus infection from latency by expression of the ORF 50 transactivator, a homolog of the EBV R protein. Virology 1998, 252, 304–312. [Google Scholar] [CrossRef] [Green Version]
- Roth, W.K.; Brandstetter, H.; Sturzl, M. Cellular and molecular features of HIV-associated Kaposi’s sarcoma. Aids 1992, 6, 895–914. [Google Scholar] [CrossRef]
- Ramos, J.C.; Sin, S.H.; Staudt, M.R.; Roy, D.; Vahrson, W.; Dezube, B.J.; Harrington, W.; Dittmer, D.P. Nuclear factor kappa B pathway associated biomarkers in AIDS defining malignancies. Int. J. Cancer 2012, 130, 2728–2733. [Google Scholar] [CrossRef] [Green Version]
- Stürzl, M.; Brandstetter, H.; Zietz, C.; Eisenburg, B.; Raivich, G.; Gearing, D.P.; Brockmeyer, N.H.; Hofschneider, P. Identification of interleukin-1 and platelet-derived growth factor-B as major mitogens for the spindle cells of Kaposi’s sarcoma: A combined in vitro and in vivo analysis. Oncogene 1995, 10, 2007. [Google Scholar]
- Miles, S.A.; Rezai, A.R.; Salazar-Gonzalez, J.F.; Vander Meyden, M.; Stevens, R.H.; Logan, D.M.; Mitsuyasu, R.T.; Taga, T.; Hirano, T.; Kishimoto, T. AIDS Kaposi sarcoma-derived cells produce and respond to interleukin 6. Proc. Natl. Acad. Sci. USA 1990, 87, 4068–4072. [Google Scholar] [CrossRef] [Green Version]
- Sciacca, F.L.; Stürzl, M.; Bussolino, F.; Sironi, M.; Brandstetter, H.; Zietz, C.; Zhou, D.; Matteucci, C.; Peri, G.; Sozzani, S. Expression of adhesion molecules, platelet-activating factor, and chemokines by Kaposi’s sarcoma cells. J. Immunol. 1994, 153, 4816–4825. [Google Scholar] [PubMed]
- Ensoli, B.; Stürzl, M.; Monini, P. Cytokine-mediated growth promotion of Kaposi’s sarcoma and primary effusion lymphoma. Semin. Cancer Biol. 2000, 10, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, M.M.; Pietrek, M.; Dittrich-Breiholz, O.; Kracht, M.; Schulz, T.P. Modulation of host gene expression by the K15 protein of Kaposi’s sarcoma-associated herpesvirus. J. Virol. 2007, 81, 42–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, J.; Damania, B. Upregulation of the TLR3 pathway by Kaposi’s sarcoma-associated herpesvirus during primary infection. J. Virol. 2008, 82, 5440–5449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couty, J.-P.; Lupu-Meiri, M.; Oron, Y.; Gershengorn, M.C. Kaposi’s sarcoma-associated herpesvirus-G protein-coupled receptor-expressing endothelial cells exhibit reduced migration and stimulated chemotaxis by chemokine inverse agonists. J. Pharmacol. Exp. Ther. 2009, 329, 1142–1147. [Google Scholar] [CrossRef]
- Mesri, E.A.; Cesarman, E.; Boshoff, C. Kaposi’s sarcoma and its associated herpesvirus. Nat. Rev. Cancer 2010, 10, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Tso, F.Y.; Kossenkov, A.V.; Lidenge, S.J.; Ngalamika, O.; Ngowi, J.R.; Mwaiselage, J.; Wickramasinghe, J.; Kwon, E.H.; West, J.T.; Lieberman, P.M. RNA-Seq of Kaposi’s sarcoma reveals alterations in glucose and lipid metabolism. PLoS Pathog. 2018, 14, e1006844. [Google Scholar] [CrossRef] [Green Version]
- Mercader, M.; Taddeo, B.; Panella, J.R.; Chandran, B.; Nickoloff, B.J.; Foreman, K.E. Induction of HHV-8 lytic cycle replication by inflammatory cytokines produced by HIV-1-infected T cells. Am. J. Pathol. 2000, 156, 1961–1971. [Google Scholar] [CrossRef] [Green Version]
- Gregory, S.M.; West, J.A.; Dillon, P.J.; Hilscher, C.; Dittmer, D.P.; Damania, B. Toll-like receptor signaling controls reactivation of KSHV from latency. Proc. Natl. Acad. Sci. USA 2009, 106, 11725–11730. [Google Scholar] [CrossRef] [Green Version]
- Riva, G.; Barozzi, P.; Torelli, G.; Luppi, M. Immunological and inflammatory features of Kaposi’s sarcoma and other Kaposi’s sarcoma-associated herpesvirus/human herpesvirus 8-associated neoplasias. Aids Rev. 2010, 12, 40–51. [Google Scholar] [PubMed]
- Ensoli, B.; Stürzl, M. Kaposi’s sarcoma: A result of the interplay among inflammatory cytokines, angiogenic factors and viral agents. Cytokine Growth Factor Rev. 1998, 9, 63–83. [Google Scholar] [CrossRef]
- Fyfe, G.; Fisher, R.I.; Rosenberg, S.A.; Sznol, M.; Parkinson, D.R.; Louie, A.C. Results of treatment of 255 patients with metastatic renal cell carcinoma who received high-dose recombinant interleukin-2 therapy. J. Clin. Oncol. 1995, 13, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Golomb, H.M.; Jacobs, A.; Fefer, A.; Ozer, H.; Thompson, J.; Portlock, C.; Ratain, M.; Golde, D.; Vardiman, J.; Burke, J. Alpha-2 interferon therapy of hairy-cell leukemia: A multicenter study of 64 patients. J. Clin. Oncol. 1986, 4, 900–905. [Google Scholar] [CrossRef]
- Groopman, J.E.; Gottlieb, M.S.; Goodman, J.; Mitsuyasu, R.T.; Conant, M.A.; Prince, H.; Fahey, J.L.; Derezin, M.; Weinstein, W.M.; Casavante, C. Recombinant alpha-2 interferon therapy for Kaposi’s sarcoma associated with the acquired immunodeficiency syndrome. Ann. Intern. Med. 1984, 100, 671–676. [Google Scholar] [CrossRef]
- Trikha, M.; Corringham, T.; Klein, B.; Rossi, J.-F. Targeted anti-interleukin-6 monoclonal antibody therapy for cancer: A review of the rationale and clinical evidence. Clin. Cancer Res. 2003, 9, 4653–4665. [Google Scholar]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Srirangan, S.; Choy, E.H. The role of interleukin 6 in the pathophysiology of rheumatoid arthritis. Ther. Adv. Musculoskelet. Dis. 2010, 2, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Moore, P.S.; Boshoff, C.; Weiss, R.A.; Chang, Y. Molecular mimicry of human cytokine and cytokine response pathway genes by KSHV. Science 1996, 274, 1739–1744. [Google Scholar] [CrossRef]
- Nicholas, J.; Ruvolo, V.R.; Burns, W.H.; Sandford, G.; Wan, X.; Ciufo, D.; Hendrickson, S.B.; Guo, H.-G.; Hayward, G.S.; Reixz, M.S. Kaposi’s sarcoma-associated human herpesvirus-8 encodes homologues of macrophage inflammatory protein-1 and interleukin-6. Nat. Med. 1997, 3, 287–292. [Google Scholar] [CrossRef]
- Sakakibara, S.; Tosato, G. Viral Interleukin-6: Role in Kaposi’s Sarcoma-Associated Herpesvirus–Associated Malignancies. J. Interferon Cytokine Res. 2011, 31, 791–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehghani, B.; Hashempour, T.; Hasanshahi, Z. Interaction of human herpesvirus 8 viral interleukin-6 with human interleukin-6 receptor using in silico approach: The potential role in HHV-8 pathogenesis. Curr. Proteom. 2020, 17, 107–116. [Google Scholar] [CrossRef]
- El-Mallawany, N.K.; Mehta, P.S.; Kamiyango, W.; Villiera, J.; Peckham-Gregory, E.C.; Kampani, C.; Krysiak, R.; Sanders, M.K.; Caro-Vegas, C.; Eason, A.B. KSHV viral load and Interleukin-6 in HIV-associated pediatric Kaposi sarcoma—Exploring the role of lytic activation in driving the unique clinical features seen in endemic regions. Int. J. Cancer 2019, 144, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Aoki, Y.; Yarchoan, R.; Wyvill, K.; Okamoto, S.-I.; Little, R.F.; Tosato, G. Detection of viral interleukin-6 in Kaposi sarcoma–associated herpesvirus–linked disorders. Blood J. Am. Soc. Hematol. 2001, 97, 2173–2176. [Google Scholar] [CrossRef]
- Lurain, K.; Polizzotto, M.N.; Aleman, K.; Bhutani, M.; Wyvill, K.M.; Gonçalves, P.H.; Ramaswami, R.; Marshall, V.A.; Miley, W.; Steinberg, S.M. Viral, immunologic, and clinical features of primary effusion lymphoma. Blood J. Am. Soc. Hematol. 2019, 133, 1753–1761. [Google Scholar]
- Jones, K.D.; Aoki, Y.; Chang, Y.; Moore, P.S.; Yarchoan, R.; Tosato, G. Involvement of interleukin-10 (IL-10) and viral IL-6 in the spontaneous growth of Kaposi’s sarcoma herpesvirus-associated infected primary effusion lymphoma cells. Blood J. Am. Soc. Hematol. 1999, 94, 2871–2879. [Google Scholar]
- Aoki, Y.; Jaffe, E.S.; Chang, Y.; Jones, K.; Teruya-Feldstein, J.; Moore, P.S.; Tosato, G. Angiogenesis and Hematopoiesis Induced by Kaposi’s Sarcoma-Associated Herpesvirus-Encoded Interleukin-6: Presented in part at the 40th Annual American Society of Hematology Meeting, December 7, 1998 (Miami Beach, FL). Blood J. Am. Soc. Hematol. 1999, 93, 4034–4043. [Google Scholar]
- Goto, H.; Kudo, E.; Kariya, R.; Taura, M.; Katano, H.; Okada, S. Targeting VEGF and interleukin-6 for controlling malignant effusion of primary effusion lymphoma. J. Cancer Res. Clin. Oncol. 2015, 141, 465–474. [Google Scholar] [CrossRef]
- Mori, Y.; Nishimoto, N.; Ohno, M.; Inagi, R.; Dhepakson, P.; Amou, K.; Yoshizaki, K.; Yamanishi, K. Human herpesvirus 8-encoded interleukin-6 homologue (viral IL-6) induces endogenous human IL-6 secretion. J. Med. Virol. 2000, 61, 332–335. [Google Scholar] [CrossRef]
- Suthaus, J.; Stuhlmann-Laeisz, C.; Tompkins, V.S.; Rosean, T.R.; Klapper, W.; Tosato, G.; Janz, S.; Scheller, J.; Rose-John, S. HHV-8–encoded viral IL-6 collaborates with mouse IL-6 in the development of multicentric Castleman disease in mice. Blood 2012, 119, 5173–5181. [Google Scholar] [CrossRef]
- Oksenhendler, E.; Carcelain, G.; Aoki, Y.; Boulanger, E.; Maillard, A.; Clauvel, J.-P.; Agbalika, F.L. High levels of human herpesvirus 8 viral load, human interleukin-6, interleukin-10, and C reactive protein correlate with exacerbation of multicentric castleman disease in HIV-infected patients. Blood J. Am. Soc. Hematol. 2000, 96, 2069–2073. [Google Scholar]
- Katsume, A.; Saito, H.; Yamada, Y.; Yorozu, K.; Ueda, O.; Akamatsu, K.-I.; Nishimoto, N.; Kishimoto, T.; Yoshizaki, K.; Ohsugi, Y. Anti-interleukin 6 (IL-6) receptor antibody suppresses Castleman’s disease like symptoms emerged in IL-6 transgenic mice. Cytokine 2002, 20, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, N.; Kanakura, Y.; Aozasa, K.; Johkoh, T.; Nakamura, M.; Nakano, S.; Nakano, N.; Ikeda, Y.; Sasaki, T.; Nishioka, K. Humanized anti–interleukin-6 receptor antibody treatment of multicentric Castleman disease. Blood 2005, 106, 2627–2632. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, H.; Kadowaki, N.; Nishikori, M.; Kitawaki, T.; Kondo, T.; Ishikawa, T.; Yoshifuji, H.; Yamakawa, N.; Imura, T.; Mimori, T. Clinical features and treatment of multicentric castleman’s disease: A retrospective study of 21 Japanese patients at a single institute. J. Clin. Exp. Hematop. 2013, 53, 69–77. [Google Scholar] [CrossRef]
- Ramaswami, R.; Lurain, K.; Peer, C.J.; Serquiña, A.; Wang, V.; Widell, A.; Goncalves, P.; Steinberg, S.M.; Marshall, V.; George, J. Tocilizumab in Patients with Symptomatic Kaposi sarcoma Herpesvirus (KSHV)-associated Multicentric Castleman disease. Blood J. 2020, 135, 2316–2319. [Google Scholar] [CrossRef]
- Van Rhee, F.; Fayad, L.; Voorhees, P.; Furman, R.; Lonial, S.; Borghaei, H.; Sokol, L.; Crawford, J.; Cornfeld, M.; Qi, M. Siltuximab, a novel anti–interleukin-6 monoclonal antibody, for Castleman’s disease. J. Clin. Oncol. 2010, 28, 3701–3708. [Google Scholar] [CrossRef]
- Van Rhee, F.; Wong, R.S.; Munshi, N.; Rossi, J.-F.; Ke, X.-Y.; Fosså, A.; Simpson, D.; Capra, M.; Liu, T.; Hsieh, R.K. Siltuximab for multicentric Castleman’s disease: A randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2014, 15, 966–974. [Google Scholar] [CrossRef]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar] [CrossRef]
- Brooks, D.G.; Trifilo, M.J.; Edelmann, K.H.; Teyton, L.; McGavern, D.B.; Oldstone, M.B. Interleukin-10 determines viral clearance or persistence in vivo. Nat. Med. 2006, 12, 1301–1309. [Google Scholar] [CrossRef]
- Kotenko, S.V.; Saccani, S.; Izotova, L.S.; Mirochnitchenko, O.V.; Pestka, S.V. Human cytomegalovirus harbors its own unique IL-10 homolog (cmvIL-10). Proc. Natl. Acad. Sci. USA 2000, 97, 1695–1700. [Google Scholar] [CrossRef] [Green Version]
- Rode, H.-J.; Bugert, J.J.; Handermann, M.; Schnitzler, P.; Kehm, R.; Janssen, W.; Delius, H.; Darai, G. Molecular characterization and determination of the coding capacity of the genome of equine herpesvirus type 2 between the genome coordinates 0.235 and 0.258 (theEcoRI DNA fragment N; 4.2 kbp). Virus Genes 1994, 9, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Haig, D.M.; Fleming, S. Immunomodulation by virulence proteins of the parapoxvirus orf virus. Vet. Immunol. Immunopathol. 1999, 72, 81–86. [Google Scholar] [CrossRef]
- Raftery, M.J.; Wieland, D.; Gronewald, S.; Kraus, A.A.; Giese, T.; Schönrich, G. Shaping phenotype, function, and survival of dendritic cells by cytomegalovirus-encoded IL-10. J. Immunol. 2004, 173, 3383–3391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Kearney, P.; Plaisance, K.; Parsons, C.H. Pivotal advance: Kaposi’s sarcoma-associated herpesvirus (KSHV)-encoded microRNA specifically induce IL-6 and IL-10 secretion by macrophages and monocytes. J. Leukoc. Biol. 2010, 87, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Machado, P.R.L.; Farias, K.J.S.; Genre, J.; Oliveira, C.J.F.; Guedes, P.M.M.; da FonsecA, B.A.L. Disseminated Kaposi’s sarcoma in patients with HIV infection correlates to high serum levels of IL-10. Viral Immunol. 2014, 27, 356–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lidenge, S.J.; Tso, F.Y.; Ngalamika, O.; Ngowi, J.R.; Mortazavi, Y.; Kwon, E.H.; Shea, D.M.; Minhas, V.; Mwaiselage, V.; Wood, C. Similar Immunological Profiles Between African Endemic and Human Immunodeficiency Virus Type 1–Associated Epidemic Kaposi Sarcoma (KS) Patients Reveal the Primary Role of KS-Associated Herpesvirus in KS Pathogenesis. J. Infect. Dis. 2019, 219, 1318–1328. [Google Scholar] [CrossRef]
- Miyazawa, M.; Noguchi, K.; Kujirai, M.; Katayama, K.; Yamagoe, S.; Sugimoto, Y. IL-10 promoter transactivation by the viral K-RTA protein involves the host-cell transcription factors, specificity proteins 1 and 3. J. Biol. Chem. 2018, 293, 662–676. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Lin, S.-F.; Gradoville, L.; Yuan, Y.; Zhu, F.; Miller, G. A viral gene that activates lytic cycle expression of Kaposi’s sarcoma-associated herpesvirus. Proc. Natl. Acad. Sci. USA 1998, 95, 10866–10871. [Google Scholar] [CrossRef] [Green Version]
- Lukac, D.M.; Kirshner, J.R.; Ganem, D. Transcriptional activation by the product of open reading frame 50 of Kaposi’s sarcoma-associated herpesvirus is required for lytic viral reactivation in B cells. J. Virol. 1999, 73, 9348–9361. [Google Scholar] [CrossRef] [Green Version]
- Staudt, M.; Dittmer, D. The Rta/Orf50 transactivator proteins of the gamma-herpesviridae. In Kaposi Sarcoma Herpesvirus: New Perspectives; Springer: Berlin/Heidelberg, Germany, 2007; pp. 71–100. [Google Scholar]
- Santarelli, R.; Gonnella, R.; Di Giovenale, G.; Cuomo, L.; Capobianchi, A.; Granato, M.; Gentile, G.; Faggioni, A.; Cirone, M. STAT3 activation by KSHV correlates with IL-10, IL-6 and IL-23 release and an autophagic block in dendritic cells. Sci. Rep. 2014, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lopes, T.R.R.; Gonçales, J.P.; Júnior, J.V.J.S.; de Lorena, V.M.B.; Toscano, A.L.C.C.; Akamatsu, S.M.; Salles, A.C.; Tozetto-Mendoza, T.R.; de Morais, V.M.S.; Coêlho, M.R.C.D. Association of IL-6, IL-10 and CXCL10 serum concentrations with visceral Kaposi’s sarcoma in people living with HIV/AIDS. Hum. Immunol. 2020, 81, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Polizzotto, M.N.; Uldrick, T.S.; Hu, D.; Yarchoan, R. Clinical manifestations of Kaposi sarcoma herpesvirus lytic activation: Multicentric Castleman disease (KSHV–MCD) and the KSHV inflammatory cytokine syndrome. Front. Microbiol. 2012, 3, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasperini, P.; Sakakibara, S.; Tosato, G. Contribution of viral and cellular cytokines to Kaposi’s sarcoma-associated herpesvirus pathogenesis. J. Leukoc. Biol. 2008, 84, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Caro-Vegas, C.; Sellers, S.; Host, K.M.; Seltzer, J.; Landis, J.; Fischer II, W.A.; Damania, B.; Dittmer, D.P. Runaway Kaposi Sarcoma-associated herpesvirus replication correlates with systemic IL-10 levels. Virology 2020, 539, 18–25. [Google Scholar] [CrossRef]
- Ejrnaes, M.; von Herrath, M.G.; Christen, U. Cure of Chronic Viral Infection and Virus-Induced Type 1 Diabetes by Neutralizing Antibodies. Clin. Dev. Immunol. 2006, 13, 810853. [Google Scholar] [CrossRef]
- Porichis, F.; Hart, M.G.; Zupkosky, J.; Barblu, L.; Kwon, D.S.; McMullen, A.; Brennan, T.; Ahmed, R.; Freeman, G.J.; Kavanagh, D.G.; et al. Differential Impact of PD-1 and/or Interleukin-10 Blockade on HIV-1-Specific CD4 T Cell and Antigen-Presenting Cell Functions. J. Virol. 2014, 88, 2508–2518. [Google Scholar] [CrossRef] [Green Version]
- Wilson, E.B.; Brooks, D.G. The role of IL-10 in regulating immunity to persistent viral infections. Curr. Top. Microbiol. Immunol. 2011, 350, 39–65. [Google Scholar]
- Rigopoulou, E.I.; Abbott, W.G.H.; Haigh, P.; Naoumov, N.V. Blocking of interleukin-10 receptor--a novel approach to stimulate T-helper cell type 1 responses to hepatitis C virus. Clin. Immunol. 2005, 117, 57–64. [Google Scholar] [CrossRef]
- Biron, C.A. Initial and innate responses to viral infections—pattern setting in immunity or disease. Curr. Opin. Microbiol. 1999, 2, 374–381. [Google Scholar] [CrossRef]
- Biron, C.A.; Nguyen, K.B.; Pien, G.C.; Cousens, L.P.; Salazar-Mather, T.P. Natural killer cells in antiviral defense: Function and regulation by innate cytokines. Annu. Rev. Immunol. 1999, 17, 189–220. [Google Scholar] [CrossRef]
- Doly, J.; Civas, A.; Navarro, S.; Uze, G. Type I interferons: Expression and signalization. Cell. Mol. Life Sci. CMLS 1998, 54, 1109–1121. [Google Scholar] [CrossRef]
- Schindler, C. Cytokines and JAK–STAT signaling. Exp. Cell Res. 1999, 253, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Schindler, C.W. Series introduction: JAK-STAT signaling in human disease. J. Clin. Investig. 2002, 109, 1133–1137. [Google Scholar] [CrossRef]
- Schindler, C.; Brutsaert, S. Interferons as a paradigm for cytokine signal transduction. Cell. Mol. Life Sci. CMLS 1999, 55, 1509–1522. [Google Scholar] [CrossRef]
- Fischl, M.A.; Finkelstein, D.M.; He, W.; Powderly, W.G.; Triozzi, P.L.; Steigbigel, R.T. A phase II study of recombinant human interferon-α2a and zidovudine in patients with AIDS-related Kaposi’s sarcoma. JAIDS J. Acquir. Immune Defic. Syndr. 1996, 11, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Krown, S.E. Interferon and other biologic agents for the treatment of Kaposi’s sarcoma. Hematol./Oncol. Clin. N. Am. 1991, 5, 311–322. [Google Scholar] [CrossRef]
- Sawyer, L.A.; Metcalf, J.A.; Zoon, K.C.; Boone, E.J.; Kovacs, J.A.; Lane, H.C.; Quinnan Jr, G.V. Effects of interferon-α in patients with aids-associated Kaposi’s sarcoma are related to blood interferon levels and dose. Cytokine 1990, 2, 247–252. [Google Scholar] [CrossRef]
- West, J.A.; Gregory, S.M.; Sivaraman, V.; Su, L.; Damania, B. Activation of plasmacytoid dendritic cells by Kaposi’s sarcoma-associated herpesvirus. J. Virol. 2011, 85, 895–904. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Hayward, G.S. The ubiquitin E3 ligase RAUL negatively regulates type i interferon through ubiquitination of the transcription factors IRF7 and IRF3. Immunity 2010, 33, 863–877. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Wang, S.E.; Hayward, G.S. The KSHV immediate-early transcription factor RTA encodes ubiquitin E3 ligase activity that targets IRF7 for proteosome-mediated degradation. Immunity 2005, 22, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.X.; King, S.M.; Smith, E.L.; Levy, D.E.; Yuan, Y. A Kaposi’s sarcoma-associated herpesviral protein inhibits virus-mediated induction of type I interferon by blocking IRF-7 phosphorylation and nuclear accumulation. Proc. Natl. Acad. Sci. USA 2002, 99, 5573–5578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damania, B.; Jacobs, S.R. The viral interferon regulatory factors of KSHV: Immunosuppressors or oncogenes? Front. Immunol. 2011, 2, 19. [Google Scholar]
- Honda, K.; Taniguchi, T. IRFs: Master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nat. Rev. Immunol. 2006, 6, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, N.; Flamand, L. Kaposi sarcoma-associated herpesvirus latency-associated nuclear antigen inhibits interferon (IFN) β expression by competing with IFN regulatory factor-3 for binding to IFNB promoter. J. Biol. Chem. 2010, 285, 7208–7221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganser, A.; Brücher, W.; Brodt, H.; Busch, W.; Brandhorst, I.; Helm, E.; Hoelzer, D. Treatment of AIDS-Related Kaposi’s Sarcoma with Recombinant γ-Interferon. Oncol. Res. Treat. 1986, 9, 163–166. [Google Scholar] [CrossRef]
- Krigel, R.L.; Odajnyk, C.M.; Laubenstein, L.J.; Ostreicher, R.; Wernz, J.; Vilcek, J.; Rubinstein, P.; Friedman-Kien, A.E. Therapeutic trial of interferon-gamma in patients with epidemic Kaposi’s sarcoma. J. Biol. Response Modif. 1985, 4, 358–364. [Google Scholar]
- Chang, J.; Renne, R.; Dittmer, D.; Ganem, D. Inflammatory cytokines and the reactivation of Kaposi’s sarcoma-associated herpesvirus lytic replication. Virology 2000, 266, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Park, M.-K.; Cho, H.; Roh, S.W.; Kim, S.-J.; Myoung, J. Cell Type-Specific Interferon-γ-mediated Antagonism of KSHV Lytic Replication. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pozharskaya, V.P.; Weakland, L.L.; Offermann, M.K. Inhibition of infectious human herpesvirus 8 production by gamma interferon and alpha interferon in BCBL-1 cells. J. Gen. Virol. 2004, 85, 2779–2787. [Google Scholar] [CrossRef]
- Bibert, S.; Wójtowicz, A.; Taffé, P.; Tarr, P.E.; Bernasconi, E.; Furrer, H.; Günthard, H.F.; Hoffmann, M.; Kaiser, L.; Osthoff, M. Interferon lambda 3/4 polymorphisms are associated with AIDS-related Kaposi’s sarcoma. Aids 2018, 32, 2759–2765. [Google Scholar] [CrossRef] [Green Version]
- Lane, H.C.; Feinberg, J.; Davey, V.; Deyton, L.; Baseler, M.; Manischewitz, J.; Masur, H.; Kovacs, J.; Herpin, B.; Walker, R. Anti-retroviral effects of interferon-α in AIDS-associated Kaposi’s sarcoma. Lancet 1988, 332, 1218–1222. [Google Scholar] [CrossRef]
- Krown, S.E.; Li, P.; Von Roenn, J.H.; Paredes, J.; Huang, J.; Testa, M.A. Efficacy of low-dose interferon with antiretroviral therapy in Kaposi’s sarcoma: A randomized phase II AIDS clinical trials group study. J. Interferon Cytokine Res. 2002, 22, 295–303. [Google Scholar] [CrossRef]
- Wu, W.; Rochford, R.; Toomey, L.; Harrington Jr, W.; Feuer, G. Inhibition of HHV-8/KSHV infected primary effusion lymphomas in NOD/SCID mice by azidothymidine and interferon-α. Leuk. Res. 2005, 29, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Rokx, C.; van der Ende, M.E.; Verbon, A.; Rijnders, B.J. Peginterferon alfa-2a for AIDS-associated Kaposi sarcoma: Experience with 10 patients. Clin. Infect. Dis. 2013, 57, 1497–1499. [Google Scholar] [CrossRef] [Green Version]
- Rouanet, I.; Lechiche, C.; Doncesco, R.; Mauboussin, J.M.; Sotto, A. Interferon therapy for Kaposi sarcoma associated with acquired immunodeficiency syndrome: Still a valid treatment option? AIDS Patient Care STDs 2013, 27, 537–538. [Google Scholar] [CrossRef]
- Kollias, G.; Douni, E.; Kassiotis, G.; Kontoyiannis, D. The function of tumour necrosis factor and receptors in models of multi-organ inflammation, rheumatoid arthritis, multiple sclerosis and inflammatory bowel disease. Ann. Rheum. Dis. 1999, 58 (Suppl. 1), I32–I39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Old, L.J. Tumor necrosis factor. Science 1985, 230, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Bazzoni, F.; Beutler, B. The tumor necrosis factor ligand and receptor families. N. Engl. J. Med. 1996, 334, 1717–1725. [Google Scholar] [CrossRef]
- Aboulafia, D.; Miles, S.A.; Saks, S.R.; Mitsuyasu, R.T. Intravenous recombinant tumor necrosis factor in the treatment of AIDS-related Kaposi’s sarcoma. JAIDS J. Acquir. Immune Defic. Syndr. 1989, 2, 54–58. [Google Scholar]
- Murakami-Mori, K.; Mori, S.; Bonavida, B.; Nakamura, S. Implication of TNF receptor-I-mediated extracellular signal-regulated kinases 1 and 2 (ERK1/2) activation in growth of AIDS-associated Kaposi’s sarcoma cells: A possible role of a novel death domain protein MADD in TNF-α-induced ERK1/2 activation in Kaposi’s sarcoma cells. J. Immunol. 1999, 162, 3672–3679. [Google Scholar]
- Oxholm, A.; Oxholm, P.; Permin, H.; Bendtzen, K. Epidermal tumour necrosis factor α and interleukin 6-like activities in AIDS-related Kaposi’s sarcoma: An immunohistological study. Apmis 1989, 97, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.A.; UAbdel-Raouf, M.; Akula, S.M. Membrane-Associated Kaposi Sarcoma-Associated Herpesvirus Glycoprotein B Promotes Cell Adhesion and Inhibits Migration of Cells via Upregulating IL-1β and TNF-α. Intervirology 2017, 60, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Gonçales, J.P.; Lopes, T.R.R.; de Lorena, V.M.B.; Aciole, M.R.; Júnior, J.V.J.S.; Tozetto-Mendoza, T.R.; Braz-Silva, P.H.; Coêlho, M.R.C.D. Association of NFκB and related-cytokines with the viral load and development of antibodies against HHV-8 in people living with HIV/AIDS. Med Microbiol. Immunol. 2020, 209, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.A.; Schattner, E.J.; Cesarman, E. Inhibition of NF-κB induces apoptosis of KSHV-infected primary effusion lymphoma cells. Blood J. Am. Soc. Hematol. 2000, 96, 2537–2542. [Google Scholar]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S. NF-ĸB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Oeckinghaus, A.; Ghosh, S. The NF-κB family of transcription factors and its regulation. Cold Spring Harbor Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Basseres, D.; Baldwin, A. Nuclear factor-κ B and inhibitor of κ B kinase pathways in oncogenic initiation and progression. Oncogene 2006, 25, 6817–6830. [Google Scholar] [CrossRef] [Green Version]
- Napetschnig, J.; Wu, H. Molecular basis of NF-κB signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef] [Green Version]
- Courtois, G.; Gilmore, T. Mutations in the NF-κ B signaling pathway: Implications for human disease. Oncogene 2006, 25, 6831–6843. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.L.; Greene, M.H.; Gershon, S.K.; Edwards, E.T.; Braun, M.M. Tumor necrosis factor antagonist therapy and lymphoma development: Twenty-six cases reported to the Food and Drug Administration. Arthritis Rheum. 2002, 46, 3151–3158. [Google Scholar] [CrossRef]
- Cohen, C.; Horster, S.; Sander, C.; Bogner, J. Kaposi’s sarcoma associated with tumour necrosis factor α neutralising therapy. Ann. Rheum. Dis. 2003, 62, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuttikat, A.; Joshi, A.; Saeed, I.; Chakravarty, K. Kaposi sarcoma in a patient with giant cell arteritis. Dermatol. Online J. 2006, 12, 16. [Google Scholar] [PubMed]
- Ursini, F.; Naty, S.; Mazzei, V.; Spagnolo, F.; Grembiale, R.D. Kaposi’s sarcoma in a psoriatic arthritis patient treated with infliximab. Int. Immunopharmacol. (Print) 2010, 10, 827–828. [Google Scholar] [CrossRef]
- Vural, S.; Gündoğdu, M.; Akay, B.N.; Korkmaz, P.; Şanli, H.; Heper, A.O.; Kundakci, N. Aggressive Kaposi’s Sarcoma Associated With Golimumab Therapy. Arch. Rheumatol. 2018, 33, 384–386. [Google Scholar] [CrossRef] [PubMed]
- Torre-Cisneros, J.; Del Castillo, J.M.; Caston, J.; Castro, M.; Perez, V.; Collantes, E. Infliximab does not activate replication of lymphotropic herpesviruses in patients with refractory rheumatoid arthritis. Rheumatology 2005, 44, 1132–1135. [Google Scholar] [CrossRef] [Green Version]
- Klausner, J.D.; VFreedman, H.; Kaplan, G. Thalidomide as an anti-TNF-α inhibitor: Implications for clinical use. Clin. Immunol. Immunopathol. 1996, 81, 219–223. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Sun, Y.; Torphy, R.J.; He, J.; Yanaga, K.; Edil, B.H.; Schulick, R.D.; Zhu, Y. Pomalidomide inhibits PD-L1 induction to promote antitumor immunity. Cancer Res. 2018, 78, 6655–6665. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Mishra, S.; Anagho, H.A.; Aisabor, A.I.; Shrestha, P.; Wang, V.; Takamatsu, Y.; Maeda, K.; Mitsuya, H.; Zeldis, J.B. Restoration of immune surface molecules in Kaposi sarcoma-associated herpes virus infected cells by lenalidomide and pomalidomide. Oncotarget 2017, 8, 50342. [Google Scholar] [CrossRef] [Green Version]
- Little, R.F.; Wyvill, K.M.; Pluda, J.M.; Welles, L.; Marshall, V.; Figg, W.D.; Newcomb, F.M.; Tosato, G.; Feigal, E.; Steinberg, S.M. Activity of thalidomide in AIDS-related Kaposi’s sarcoma. J. Clin. Oncol. 2000, 18, 2593–2602. [Google Scholar] [CrossRef]
- Polizzotto, M.N.; Uldrick, T.S.; Wyvill, K.M.; Aleman, K.; Peer, C.J.; Bevans, M.; Sereti, I.; Maldarelli, F.; Whitby, D.; Marshall, V. Pomalidomide for symptomatic Kaposi’s sarcoma in people with and without HIV infection: A phase I/II study. J. Clin. Oncol. 2016, 34, 4125. [Google Scholar] [CrossRef]
- Stern, A.S.; Podlaski, F.J.; Hulmes, J.D.; Pan, Y.; Quinn, P.M.; Wolitzky, A.; Familletti, P.C.; Stremlo, D.L.; Truitt, T.; Chizzonite, R. Purification to homogeneity and partial characterization of cytotoxic lymphocyte maturation factor from human B-lymphoblastoid cells. Proc. Natl. Acad. Sci. USA 1990, 87, 6808–6812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakóbisiak, M.; Lasek, W.; Gołąb, J. Natural mechanisms protecting against cancer. Immunol. Lett. 2003, 90, 103–122. [Google Scholar] [CrossRef]
- Cesarman, E.; Nador, R.G.; Bai, F.; Bohenzky, R.A.; Russo, J.J.; Moore, P.S.; Chang, Y.; Knowles, D.M. Kaposi’s sarcoma-associated herpesvirus contains G protein-coupled receptor and cyclin D homologs which are expressed in Kaposi’s sarcoma and malignant lymphoma. J. Virol. 1996, 70, 8218–8223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geras-Raaka, E.; Varma, A.; Ho, H.; Clark-Lewis, I.; Gershengorn, M.C. Human Interferon-γ–inducible Protein 10 (IP-10) Inhibits Constitutive Signaling of Kaposi’s Sarcoma–associated Herpesvirus G Protein–coupled Receptor. J. Exp. Med. 1998, 188, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sodhi, A.; Montaner, S.; Patel, V.; Zohar, M.; Bais, C.; Mesri, E.A.; Gutkind, J.S. The Kaposi’s sarcoma-associated herpes virus G protein-coupled receptor up-regulates vascular endothelial growth factor expression and secretion through mitogen-activated protein kinase and p38 pathways acting on hypoxia-inducible factor 1α. Cancer Res. 2000, 60, 4873–4880. [Google Scholar] [PubMed]
- Bais, C.; Santomasso, B.; Coso, O.; Arvanitakis, L.; Raaka, E.G.; Gutkind, J.S.; Asch, A.S.; Cesarman, E.; Gerhengorn, M.C.; Mesri, E.A. G-protein-coupled receptor of Kaposi’s sarcoma-associated herpesvirus is a viral oncogene and angiogenesis activator. Nature 1998, 391, 86–89. [Google Scholar] [CrossRef]
- Nakamura, S.; Murakami-Mori, K.; Rao, N.; Weich, H.A.; Rajeev, B. Vascular endothelial growth factor is a potent angiogenic factor in AIDS-associated Kaposi’s sarcoma-derived spindle cells. J. Immunol. 1997, 158, 4992–5001. [Google Scholar]
- Little, R.F.; Pluda, J.M.; Wyvill, K.M.; Rodriguez-Chavez, I.R.; Tosato, G.; Catanzaro, A.T.; Steinberg, S.M.; Yarchoan, R. Activity of subcutaneous interleukin-12 in AIDS-related Kaposi sarcoma. Blood 2006, 107, 4650–4657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, R.F.; Aleman, K.; Kumar, P.; Wyvill, K.M.; Pluda, J.M.; Read-Connole, E.; Wang, V.; Pittaluga, S.; Catanzaro, A.T.; Steinberg, S.M. Phase 2 study of pegylated liposomal doxorubicin in combination with interleukin-12 for AIDS-related Kaposi sarcoma. Blood J. Am. Soc. Hematol. 2007, 110, 4165–4171. [Google Scholar] [CrossRef] [Green Version]
- Sharifi, J.; Khawli, L.A.; Hu, P.; King, S.; Epstein, A.L. Characterization of a phage display-derived human monoclonal antibody (NHS76) counterpart to chimeric TNT-1 directed against necrotic regions of solid tumors. Hybrid. Hybridomics 2001, 20, 305–312. [Google Scholar] [CrossRef]
- Neipel, F.; Albrecht, J.-C.; Fleckenstein, B. Cell-homologous genes in the Kaposi’s sarcoma-associated rhadinovirus human herpesvirus 8: Determinants of its pathogenicity? J. Virol. 1997, 71, 4187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, J.J.; Bohenzky, R.A.; Chien, M.-C.; Chen, J.; Yan, M.; Maddalena, D.; Parry, J.P.; Peruzzi, D.; Edelman, I.S.; Chang, Y. Nucleotide sequence of the Kaposi sarcoma-associated herpesvirus (HHV8). Proc. Natl. Acad. Sci. USA 1996, 93, 14862–14867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenner, R.G.; Albà, M.M.; Boshoff, C.; Kellam, P. Kaposi’s sarcoma-associated herpesvirus latent and lytic gene expression as revealed by DNA arrays. J. Virol. 2001, 75, 891–902. [Google Scholar] [CrossRef] [Green Version]
- Paulose-Murphy, M.; Ha, N.-K.; Xiang, C.; Chen, Y.; Gillim, L.; Yarchoan, R.; Meltzer, P.; Bittner, M.; Trent, J.; Zeichner, S. Transcription program of human herpesvirus 8 (Kaposi’s sarcoma-associated herpesvirus). J. Virol. 2001, 75, 4843–4853. [Google Scholar] [CrossRef] [Green Version]
- Dairaghi, D.J.; Fan, R.A.; McMaster, B.E.; Hanley, M.R.; Schall, T.J. HHV8-encoded vMIP-I selectively engages chemokine receptor CCR8 Agonist and antagonist profiles of viral chemokines. J. Biol. Chem. 1999, 274, 21569–21574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endres, M.J.; Garlisi, C.G.; Xiao, H.; Shan, L.; Hedrick, J.A. The Kaposi’s sarcoma–related herpesvirus (KSHV)-encoded chemokine vMIP-I is a specific agonist for the CC chemokine receptor (CCR) 8. J. Exp. Med. 1999, 189, 1993–1998. [Google Scholar] [CrossRef] [Green Version]
- Lüttichau, H.R.; Johnsen, A.H.; Jurlander, J.; Rosenkilde, M.M.; Schwartz, T.W. Kaposi sarcoma-associated herpes virus targets the lymphotactin receptor with both a broad spectrum antagonist vCCL2 and a highly selective and potent agonist vCCL3. J. Biol. Chem. 2007, 282, 17794–17805. [Google Scholar] [CrossRef] [Green Version]
- Haque, N.S.; Fallon, J.T.; Taubman, M.B.; Harpel, P.C. The chemokine receptor CCR8 mediates human endothelial cell chemotaxis induced by I-309 and Kaposi sarcoma herpesvirus-encoded vMIP-I and by lipoprotein (a)-stimulated endothelial cell conditioned medium. Blood J. Am. Soc. Hematol. 2001, 97, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Okruzhnov, Y.; Li, H.; Nicholas, J. Human herpesvirus 8 (HHV-8)-encoded cytokines induce expression of and autocrine signaling by vascular endothelial growth factor (VEGF) in HHV-8-infected primary-effusion lymphoma cell lines and mediate VEGF-independent antiapoptotic effects. J. Virol. 2001, 75, 10933–10940. [Google Scholar] [CrossRef] [Green Version]
- Szpakowska, M.; Chevigné, A. vCCL2/vMIP-II, the viral master KEYmokine. J. Leukoc. Biol. 2016, 99, 893–900. [Google Scholar] [CrossRef]
- Kledal, T.N.; Rosenkilde, M.M.; Coulin, F.; Simmons, G.; Johnsen, A.H.; Alouani, S.; Power, C.A.; Lüttichau, H.R.; Gerstoft, J.; Clapham, P.R. A broad-spectrum chemokine antagonist encoded by Kaposi’s sarcoma-associated herpesvirus. Science 1997, 277, 1656–1659. [Google Scholar] [CrossRef] [PubMed]
- Lüttichau, H.R.; Lewis, I.C.; Gerstoft, J.; Schwartz, T. The herpesvirus 8-encoded chemokine vMIP-II, but not the poxvirus-encoded chemokine MC148, inhibits the CCR10 receptor. Eur. J. Immunol. 2001, 31, 1217–1220. [Google Scholar] [CrossRef]
- Yamin, R.; Kaynan, N.S.; Glasner, A.; Vitenshtein, A.; Tsukerman, P.; Bauman, Y.; Ophir, Y.; Elias, S.; Bar-On, Y.; Gur, C. The viral KSHV chemokine vMIP-II inhibits the migration of Naive and activated human NK cells by antagonizing two distinct chemokine receptors. PLoS Pathog. 2013, 9, e1003568. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.B.; Nicholas, J. Autocrine and paracrine promotion of cell survival and virus replication by human herpesvirus 8 chemokines. J. Virol. 2008, 82, 6501–6513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Bacon, K.B.; Li, L.; Garcia, G.E.; Xia, Y.; Lo, D.; Thompson, D.A.; Siani, M.A.; Yamamoto, T.; Harrison, J.K. In vivo inhibition of CC and CX3C chemokine–induced leukocyte infiltration and attenuation of glomerulonephritis in Wistar-Kyoto (WKY) rats by vMIP-II. J. Exp. Med. 1998, 188, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Sozzani, S.; Luini, W.; Bianchi, G.; Allavena, P.; Wells, T.; Napolitano, M.; Bernardini, G.; Vecchi, A.; D’Ambrosio, D.; Mazzeo, D. The viral chemokine macrophage inflammatory protein-II is a selective Th2 chemoattractant. Blood J. Am. Soc. Hematol. 1998, 92, 4036–4039. [Google Scholar]
- Nakano, K.; Isegawa, Y.; Zou, P.; Tadagaki, K.; Inagi, R.; Yamanishi, K. Kaposi’s sarcoma-associated herpesvirus (KSHV)-encoded vMIP-I and vMIP-II induce signal transduction and chemotaxis in monocytic cells. Arch. Virol. 2003, 148, 871–890. [Google Scholar] [CrossRef]
- Ghirnikar, R.; Lee, Y.; Eng, L. Chemokine antagonist infusion attenuates cellular infiltration following spinal cord contusion injury in rat. J. Neurosci. Res. 2000, 59, 63–73. [Google Scholar] [CrossRef]
- Cherqui, S.; Kingdon, K.M.; Thorpe, C.; Kurian, S.M.; Salomon, D.R. Lentiviral gene delivery of vMIP-II to transplanted endothelial cells and endothelial progenitors is proangiogenic in vivo. Mol. Ther. 2007, 15, 1264–1272. [Google Scholar] [CrossRef]
- DeBruyne, L.; Li, K.; Bishop, D.; Bromberg, J. Gene transfer of virally encoded chemokine antagonists vMIP-II and MC148 prolongs cardiac allograft survival and inhibits donor-specific immunity. Gene Ther. 2000, 7, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Pillai, R.G.; Beutelspacher, S.C.; Larkin, D.F.P.; George, A.J. Expression of the chemokine antagonist vMIP II using a non-viral vector can prolong corneal allograft survival. Transplantation 2008, 85, 1640–1647. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Fan, X.; Zhou, N.; Hiraoka, M.; Luo, J.; Kaji, H.; Huang, Z. Structure−function study and anti-HIV activity of synthetic peptide analogues derived from viral chemokine vMIP-II. Biochemistry 2000, 39, 13545–13550. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Luo, Z.; Luo, J.; Hall, J.W.; Huang, Z. A novel peptide antagonist of CXCR4 derived from the N-terminus of viral chemokine vMIP-II. Biochemistry 2000, 39, 3782–3787. [Google Scholar] [CrossRef] [PubMed]
- Stine, J.T.; Wood, C.; Hill, M.; Epp, A.; Raport, C.J.; Schweickart, V.L.; Endo, Y.; Sasaki, T.; Simmons, G.; Boshoff, C. KSHV-encoded CC chemokine vMIP-III is a CCR4 agonist, stimulates angiogenesis, and selectively chemoattracts TH2 cells. Blood J. Am. Soc. Hematol. 2000, 95, 1151–1157. [Google Scholar] [CrossRef]
- Rosenkilde, M.M.; Kledal, T.N.; Bräuner-Osborne, H.; Schwartz, T.W. Agonists and inverse agonists for the herpesvirus 8-encoded constitutively active seven-transmembrane oncogene product, ORF-74. J. Biol. Chem. 1999, 274, 956–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvanitakis, L.; Geras-Raaka, E.; Varma, A.; Gershengorn, M.C.; Cesarman, E. Human herpesvirus KSHV encodes a constitutively active G-protein-coupled receptor linked to cell proliferation. Nature 1997, 385, 347–350. [Google Scholar] [CrossRef]
- De Munnik, S.M.; Kooistra, A.J.; Van Offenbeek, J.; Nijmeijer, S.; de Graaf, C.; Smit, M.J.; Leurs, R.; Vischer, H.F. The viral G protein-coupled receptor ORF74 hijacks β-arrestins for endocytic trafficking in response to human chemokines. PLoS ONE 2015, 10, e0124486. [Google Scholar] [CrossRef] [Green Version]
- Gershengorn, M.C.; Geras-Raaka, E.; Varma, A.; Clark-Lewis, I. Chemokines activate Kaposi’s sarcoma-associated herpesvirus G protein-coupled receptor in mammalian cells in culture. J. Clin. Investig. 1998, 102, 1469–1472. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Cao, Y.; Jiang, W.; Zabaleta, J.; Liu, Z.; Qiao, J.; Qin, Z. KSHV co-infection down-regulates HPV16 E6 and E7 from cervical cancer cells. Oncotarget 2017, 8, 35792. [Google Scholar] [CrossRef]
- Qian, B.-Z.; Li, J.; Zhang, H.; Kitamura, T.; Zhang, J.; Campion, L.R.; Kaiser, E.A.; Snyder, L.A.; Pollard, J.W. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 2011, 475, 222–225. [Google Scholar] [CrossRef] [Green Version]
- Wolf, M.J.; Hoos, A.; Bauer, J.; Boettcher, S.; Knust, M.; Weber, A.; Simonavicius, N.; Schneider, C.; Lang, M.; Stürzl, M. Endothelial CCR2 signaling induced by colon carcinoma cells enables extravasation via the JAK2-Stat5 and p38MAPK pathway. Cancer Cell 2012, 22, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caselli, E.; Benedetti, S.; Grigolato, J.; Caruso, A.; Di Luca, D. Activating transcription factor 4 (ATF4) is upregulated by human herpesvirus 8 infection, increases virus replication and promotes proangiogenic properties. Arch. Virol. 2012, 157, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Giffin, L.; Yan, F.; Major, M.B.; Damania, B. Modulation of Kaposi’s sarcoma-associated herpesvirus interleukin-6 function by hypoxia-upregulated protein 1. J. Virol. 2014, 88, 9429–9441. [Google Scholar] [CrossRef] [Green Version]
- Desnoyer, A.; Dupin, N.; Assoumou, L.; Carlotti, A.; Gaudin, F.; Deback, C.; Peytavin, G.; Marcelin, A.; Boué, F.; Balabanian, K. Expression pattern of the CXCL 12/CXCR 4–CXCR 7 trio in Kaposi sarcoma skin lesions. Br. J. Dermatol. 2016, 175, 1251–1262. [Google Scholar] [CrossRef]
- Miyake, M.; Lawton, A.; Goodison, S.; Urquidi, V.; Rosser, C.J. Chemokine (CXC motif) ligand 1 (CXCL1) protein expression is increased in high-grade prostate cancer. Pathol. Res. Pract. 2014, 210, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Lawton, A.; Goodison, S.; Urquidi, V.; Gomes-Giacoia, E.; Zhang, G.; Ross, S.; Kim, J.; Rosser, C.J. Chemokine (CXC) ligand 1 (CXCL1) protein expression is increased in aggressive bladder cancers. BMC Cancer 2013, 13, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, M.; Furuya, H.; Onishi, S.; Hokutan, K.; Anai, S.; Chan, O.; Shi, S.; Fujimoto, K.; Goodison, S.; Cai, W. Monoclonal antibody against CXCL1 (HL2401) as a novel agent in suppressing IL6 expression and tumoral growth. Theranostics 2019, 9, 853. [Google Scholar] [CrossRef]
- Kalbasi, A.; Komar, C.; Tooker, G.M.; Liu, M.; Lee, J.W.; Gladney, W.L.; Ben-Josef, E.; Beatty, G.L. Tumor-derived CCL2 mediates resistance to radiotherapy in pancreatic ductal adenocarcinoma. Clin. Cancer Res. 2017, 23, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Bonapace, L.; Coissieux, M.-M.; Wyckoff, J.; Mertz, K.D.; Varga, Z.; Junt, T.; Bentires-Alj, M. Cessation of CCL2 inhibition accelerates breast cancer metastasis by promoting angiogenesis. Nature 2014, 515, 130–133. [Google Scholar] [CrossRef]
- Binder, P.S.; Cullinan, D.; Nywening, T.; Wilkinson-Ryan, I.; Belt, B.; Goedegebuure, P.; Powell, M.A.; Mutch, D.G.; Hawkins, W. CCR2 blockade alters the tumor microenvironment immune infiltrate and enhances anti-tumor activity in ovarian cancer. Gynecol. Oncol. 2017, 145, 36. [Google Scholar] [CrossRef]



{kind=link}
| Cytokine | KSHV Disease | Drug | Dose | Study Type | Summary of the Result |
|---|---|---|---|---|---|
| IL-6 | MCD | Tocilizumab | 8 mg/kg MRA biweekly | Open-label trial in 28 patients for 16 weeks [43] | Alleviated the inflammatory symptoms and biochemical abnormalities |
| 8 mg/kg every 2 weeks | Ongoing Phase II clinical trial (NCT01441063). | ||||
| Tocilizumab Zidovudine Valganciclovir | 8 mg/kg IV on day 1 of 14-day cycles for a maximum of 6 cycles 600mg every 6 h 900mg every 12 h | Open label, single center pilot study [45] | One patient had complete clinical benefit response and four patients had partial clinical benefit responses on tocilizumab alone. Three patients who had AZT/VGC in combination with tocilizumab, two had partial clinical benefit responses and one had complete clinical benefit response | ||
| Siltuximab | Cohorts 1 to 5 enrolled in sequential order. 2-h infusion at 3 mg/kg every 2 weeks, 6 mg/kg every 2 weeks, 12 mg/kg every 3 weeks, 6 mg/kg weekly, and 12 mg/kg every 2 weeks, respectively. Cohort 6, 1-h infusion at 12 mg/kg every 3 weeks. Cohort 7, 9 mg/kg every 3 weeks. | Phase I, open-label, dose-finding and seven-cohort study in 23 patients [46] | 18 out of 23 patients had clinical benefits response and 12 patients showed objective tumor response. 11 patients treated with the highest dose of 12 mg/kg had clinical benefit response and 8 patients showed objective tumor response | ||
| 11 mg/kg intravenous infusion every 3 weeks | Randomized, double-blind, placebo-controlled trial in 79 patients [47]. | 18 of 53 had tumor and symptomatic responses | |||
| IFNa | AIDS-KS | IFNa | Starting dose 35×106 Reduced to 5–10×106 | Open therapeutic trial [92] | Anti-viral effects Anti-tumor response in 38% of patients |
| AIDS-KS | IFNa with didanosine | 1 million (low dose) or 10 million (intermediate dose) IU per day by subcutaneous (s.c.) injection. | randomized phase II clinical trials [93] | 40% tumor response | |
| TNFa | AIDS-KS | Thalidomide | 100 mg/day for 12 months | Phase II dose-escalation study [120] | Improved the clinical manifestation 8 out of 17 patients achieved partial response and 2 patients had stable conditions |
| AIDS-KS and KS | Pomalidomide | 5 mg/day for 21/28 days | phase I/II study in 22 KS patients with and without HIV infection. [121] | 16 patients. Achieved objective tumor response and 9 out of 15 HIV infected patients. Achieved objective response. | |
| IL-12 | AIDS-KS | L12 | 100, 300, 500 and 625 ng/kg | Phase 1 pilot study [129] | 17 had a complete or partial KS tumor response (61%) with three highest does. |
| AIDS-KS | IL12 alone and in combination with pegylated liposomal | 300 ng/kg subcutaneously twice weekly for six 3-week cycles, followed by 500 ng/kg subcutaneous IL-12 twice weekly | Phase II clinical trial [130] | 83.3% complete or partial KS tumor response. | |
| Advanced KS | NHS-IL12 | Ongoing Phase I/II clinical trial (NCT04303117) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alomari, N.; Totonchy, J. Cytokine-Targeted Therapeutics for KSHV-Associated Disease. Viruses 2020, 12, 1097. https://doi.org/10.3390/v12101097
Alomari N, Totonchy J. Cytokine-Targeted Therapeutics for KSHV-Associated Disease. Viruses. 2020; 12(10):1097. https://doi.org/10.3390/v12101097
Chicago/Turabian StyleAlomari, Nedaa, and Jennifer Totonchy. 2020. "Cytokine-Targeted Therapeutics for KSHV-Associated Disease" Viruses 12, no. 10: 1097. https://doi.org/10.3390/v12101097
APA StyleAlomari, N., & Totonchy, J. (2020). Cytokine-Targeted Therapeutics for KSHV-Associated Disease. Viruses, 12(10), 1097. https://doi.org/10.3390/v12101097