

Abstract
The axon initial segment (AIS) and nodes of Ranvier are the sites of action potential initiation and regeneration in axons. Although the basic molecular architectures of AIS and nodes, characterized by dense clusters of Na+ and K+ channels, are similar, firing patterns vary among cell types. Neuronal firing patterns are established by the collective activity of voltage-gated ion channels and can be modulated through interaction with auxiliary subunits. Here, we report the neuronal expression pattern and subcellular localization of Navβ4, the modulatory Na+ channel subunit thought to underlie resurgent Na+ current. Immunostaining of rat tissues revealed that Navβ4 is strongly enriched at the AIS of a select set of neuron types, including many characterized by high-frequency firing, and at nodes of Ranvier in the PNS and some nodes in the CNS. By introducing full-length and mutant GFP-tagged Navβ4 into cultured neurons, we determined that the AIS and nodal localization of Navβ4 depends on its direct interaction with Na+ channel α subunits through an extracellular disulfide bond. Based on these results, we propose that differences in the specific composition of the Na+ channel complexes enriched at the AIS and nodes contribute to the diverse physiologies observed among cell types.
Introduction
High-density clusters of voltage-gated Na+ channels decorate the axon initial segment (AIS) (Kole et al., 2008; Lorincz and Nusser, 2010). The activity of these AIS Na+ channels regulates the probability of action potential (AP) initiation and contributes to overall neuronal excitability (Khaliq and Raman, 2006; Palmer and Stuart, 2006; Kole and Stuart, 2008). Na+ channels undergo multiple posttranslational modifications, including glycosylation and phosphorylation, that influence their activation and inactivation kinetics. Na+ channels are also influenced by interaction with auxiliary subunits (Isom et al., 1992). Na+ channels comprise a pore-forming α subunit and modulatory β subunits. Four different Navβ subunits, termed Navβ1–Navβ4, are expressed in the nervous system. Of these, Navβ4 is uniquely associated with resurgent Na+ current (INaR), and its expression has been postulated to facilitate high-frequency AP generation during repetitive firing (Khaliq et al., 2003; Grieco et al., 2005).
Resurgent Na+ current is a transient, voltage-dependent influx of Na+ ions that occurs during repolarization of the membrane after strong depolarizations (Raman and Bean, 1997). Neurons expressing INaR are characterized by noncanonically short Na+ channel refractory periods and heightened open-channel probability at subthreshold voltages. After depolarization, INaR-carrying channels are prevented from entering classical inactivation by the rapid insertion of an open-channel blocker (Raman and Bean, 2001). As repolarization proceeds, the pore blocker disengages from the α subunit allowing an influx of Na+ ions as channels reopen before inactivating or reactivating. The C-terminal tail of Navβ4 has been proposed as the open-channel blocker underlying resurgent kinetics (Grieco et al., 2005). Knockdown of Navβ4 in cerebellar granule cells results in the loss of INaR and a decrease in repetitive firing (Bant and Raman, 2010). Inclusion of the putative blocking peptide in the recording pipette is sufficient to restore both properties in shRNA-treated cells, suggesting that Navβ4 is indeed the endogenous open-channel blocker regulating INaR. Although Navβ4 is most notably associated with INaR, its expression also increases persistent Na+ current (INaP) (Aman et al., 2009), a current implicated in neocortical pyramidal neuron burst firing (Kole, 2011).
Despite recent studies implicating Navβ4 as an important modulator of INaR and INaP (Grieco et al., 2005; Aman et al., 2009; Bant and Raman, 2010), the subcellular localization of Navβ4 remains poorly characterized (Yu et al., 2003). Here, we report that Navβ4 immunoreactivity is enriched at the AIS of cell types known to have INaR, including cerebellar Purkinje neurons and subthalamic neurons, among others (Bean 2007). We also observed layer-specific expression and AIS enrichment of Navβ4 in cortical pyramidal neurons. Furthermore, we found Navβ4 enriched at subsets of CNS and PNS nodes. Through mutational analysis, we identified the molecular mechanisms that regulate Navβ4 recruitment to the AIS and nodes. These results provide molecular evidence that the expression pattern, subcellular localization, and molecular interactions of Navβ4 are consistent with its proposed physiological role in regulating repetitive firing in multiple neuron types.
Materials and Methods
Animals.
Adult and timed-pregnant Sprague Dawley rats were purchased from Harlan Sprague Dawley. Animals were housed and maintained in Baylor College of Medicine's Center for Comparative Medicine, compliant with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Antibodies.
The following primary antibodies were used: mouse anti-α-Navβ4 (N168/6 generated against a fusion protein including amino acids 184–228; NeuroMab), rabbit α-Navβ4 F3888 (generated against a peptide, CKKLITFILKKTREK), rat α-GFP (04404-84; Nacalai Tesque), rabbit α-V5 (V8137; Sigma), mouse α-V5 (SV5-Pk1; AbD Serotec), rabbit α-Nav1.6 (Rasband et al., 2003), rabbit α-βIV spectrin (Yang et al., 2004), mouse α-Kv1.2 (K14/16; NeuroMab), chicken α-MAP2 (EnCor Biotechnology), mouse α-ankyrin G (ankG) (N106/36; NeuroMab), mouse α-pan Navα (K58/35; Sigma), rabbit α-parvalbumin (PV) (NB120-11427; Novus Biologicals), rabbit α-calbindin (CB) (C2724; Sigma), mouse α-CB (AF2E5; Abcam), rabbit α-Kv1.2 (Rhodes et al., 1995), rat α-MBP (MAB386; Millipore), rabbit α-GFP (A11122; Invitrogen), mouse α-GFP (11E5; Invitrogen), mouse α-His tag (05-531; Millipore), rabbit α-GAPDH (G9545; Sigma), and mouse α-actin (MAB1501; Millipore). The following secondary antibodies were used: goat α-rat 488 (A11006; Invitrogen), goat α-mouse XA 594 (A11032; Invitrogen), goat α-mouse IgG1 488 (115-515-205; Jackson ImmunoResearch), goat α-mouse IgG2a 594 (115-515-206; Jackson ImmunoResearch), goat α-mouse IgG2b 594 (115-515-207; Jackson ImmunoResearch), goat α-rabbit 594 (A11037; Invitrogen), goat α-rabbit 488 (A11034; Invitrogen), goat α-mouse XA 350 (A21120; Invitrogen), goat α-chicken AMCA (103-155-155; Jackson ImmunoResearch), goat α-mouse HRP (A10551; Invitrogen), and goat α-rabbit HRP (G21234; Invitrogen) at 1:1000. Hoechst staining was used to identify nuclei in some experiments.
DNA and shRNA constructs.
Navβ4 was cloned from purified rat cDNA and inserted into vector pEGFP-N1. To generate the terminal truncation mutants, we subcloned the portion of interest out of the full-length (FL) Navβ4 pEGFP–N1 construct and reinserted the product into empty pEGFP–N1 vector. Single point mutations were introduced into the FL, GFP-tagged Navβ4 DNA by site-directed mutagenesis using the QuikChange Lightning kit (Agilent Technologies). Terminal truncations and single-point mutations of Navβ4 DNA were verified by sequencing (GENEWIZ) and analysis using ApE (A Plasmid Editor) software. The efficacies of the Na+ channel (Nav1.x), neurofascin-186, and βIV spectrin shRNA expression plasmids were reported previously by Hedstrom et al. (2007).
Transfection and immunostaining of COS-1 cells.
COS-1 cells were transfected with GFP-tagged Navβ4, V5/His-tagged Navβ1, or V5/His-tagged Navβ2 mammalian expression vectors by lipid-based transfection. The Navβ1 and Navβ2 expression constructs were a gift from Dr. Doo Yeon Kim (Harvard Medical School, Boston, MA). COS-1 cells were seeded at 10% confluence and grown at 37°C in DMEM containing 10% fetal calf serum (FC-III), 1% Glutamax, and 1% penicillin–streptomycin (all from Invitrogen). Cells were transfected at ∼60% confluency. Lipofectamine 2000 and cDNA were incubated separately in serum-free Opti-MEM and then combined and incubated 30 min at room temperature. The reaction mixture (2 μl Lipofectamine 2000/μg cDNA) was then added to the COS-1 cells, which were allowed to grow 48 h in the presence of the transfection reagents and then either fixed for immunostaining or scraped and suspended in reducing sample buffer for Western blotting.
Hippocampal neuron cultures.
Hippocampal neurons were cultured essentially as in the study by Kaech and Banker (2006), in the absence of the glial feeder layer. In brief, hippocampi were isolated from the brains of E18 rat embryos, trypsin digested (0.25% trypsin in HBSS), and dissociated by trituration before plating on poly-l-lysine- and laminin-coated coverslips. Neuronal growth media (97% Neurobasal, 2% B-27 supplement, and 1% Glutamax; Invitrogen) was changed completely 4 h after plating and then supplemented, in part, with new media every 4 d.
Transfection of cultured hippocampal neurons.
Hippocampal neurons were transfected either at the time of plating (DIV0) or on DIV11. For DIV0, immediately after dissection and dissociation, neurons were nucleofected with shRNA or cDNA expression plasmids using the Neon system (Invitrogen). The cells were dissociated, centrifuged, and resuspended in buffer T at a density of 24,000 cells/μl. A single 1400 mV pulse was delivered over 20 ms to electroporate the cell membrane and introduce cDNA and/or shRNA plasmids. Neurons were plated on poly-l-lysine- and laminin-coated coverslips at a final density of 250 cells/mm2. Media was completely replaced 6 h after transfection. Hippocampal neurons were maintained from 1 to 3 weeks as indicated in primary neuron cultures methods section. For DIV11, Lipofectamine 2000 reagent (Invitrogen) was used to transfect DIV11 hippocampal neurons with Navβ4 construct cDNA. Lipofectamine and cDNA were incubated separately in Neurobasal medium and then combined. After 30 min, the reaction containing 2 μl Lipofectamine 2000/μg DNA was added to the neurons. Media was completely replaced after 4 h incubation with the transfection reagent. Cells were allowed to mature 3 d and then fixed for additional processing on DIV14.
Myelinating cocultures.
Myelinating cocultures were performed as described by Susuki et al. (2011). In brief, dorsal root ganglia (DRGs) were isolated from E16 rat embryos of either sex, dissociated, and plated on coverslips coated with Matrigel (BD Biosciences). Non-neuronal cells were eliminated by two rounds of incubation with anti-mitotic agents. The purified DRG were transfected with FL Navβ4–GFP, Navβ4ΔC–GFP, Navβ4ΔN–GFP, or Navβ4–C28A–GFP expression constructs on DIV9 using Lipofectamine LTX and PLUS reagents (Invitrogen). Purified rat Schwann cells (cultured as in the study by Susuki et al., 2011) were added at 10 DIV. Ascorbic acid was added at 15 DIV to induce myelination, and the cocultures were allowed to mature for 17 additional days. Cells were fixed in ice-cold, 4% paraformaldehyde for 20 min on DIV32 and then immunostained as described.
Western blots.
Rat brains were rapidly extracted, and the regions were isolated in ice-cold cutting solution containing the following (in mm): 110 sucrose, 60 NaCl, 3 KCl, 1.25 NaH2PO4, 28 NaHCO3, 0.5 CaCl2, and 7 MgCl2. The tissue was immediately snap frozen on dry ice. Frozen tissue was placed in ice-cold homogenization buffer containing the following: 40 mm HEPES, pH 7.5, 150 mm NaCl, 10 mm pyrophosphate, 10 mm glyceropyrophosphate, 1 mm EDTA, and 0.3% CHAPS, supplemented with protease inhibitor cocktail (Sigma) and the phosphatase inhibitors Na+-fluoride (10 μm) and Na+-orthovandate (10 μm). Membrane proteins were isolated by centrifugation. After protein quantification by BCA assay (Thermo Fisher Scientific), protein samples were diluted in either reducing or non-reducing SDS sample buffer (as indicated) and denatured at 95°C for 3 min. Subsequently, 10 or 50 μg protein/lane was loaded onto 7.5% SDS-PAGE gels, resolved, transferred onto nitrocellulose membranes, and probed overnight with the indicated primary antibodies in 5% milk, 0.2% Tween 20 TBS using standard techniques. HRP-conjugated secondary antibodies (1:1000) (Invitrogen) were used to visualize the primary antibodies. All Western blots were developed using enhanced chemiluminescence. Peroxidase activity was detected using the SuperSignal West Pico or Femto Chemiluminescence substrates (Thermo Fisher Scientific). Images were acquired on a luminescence image analyzer (GE Healthcare) through LAS4000 (GE Healthcare) software.
Immunostaining.
For whole brain, optic nerve, spinal cord, and sciatic nerve, rats were killed by an overdose of isoflurane followed by exsanguination. Nervous system tissue was dissected rapidly. Brains were drop fixed in 4% PFA and 0.1 m PB for 1.5 h and equilibrated in 20% sucrose and 0.1 m PB over 48 h. Nerves were fixed in 4% PFA and 0.1 m PB for 30 min and equilibrated in 20% sucrose and 0.1 m PB for at least 24 h. Afterward, 30 μm brain/cerebellum or 12 μm longitudinal slices (optic nerve, sciatic nerve, and dorsal and ventral roots) were cut on a microtome and washed in 0.1 m PB. Slices were blocked in 10% normal goat serum and 0.1 m PB containing 0.3% Triton X-100 (PBTGS). Tissue was incubated overnight at 4°C in primary antibodies diluted in PBTGS. Primary antibodies were removed by washing the tissue three times for 5 min with PBTGS. Secondary antibodies were diluted in PBTGS and applied for 1 h at room temperature to visualize primary antibodies. Excess secondary antibodies were removed by consecutive 5 min washes with each of PBTGS, 0.1 m PB, and 0.05 m PB. Slices were mounted on 1% gelatin-coated coverslips. For cultured neurons, cells were fixed in ice-cold 4% PFA in 0.1 m PB at 4°C for 20 min. The PFA was removed by washing three times for 5 min with ice-cold 1× Dulbecco's phosphate buffered saline. Cells were blocked and permeated in PBTGS for 30 min. Primary antibodies, diluted in PBTGS, were applied overnight at 4°C. The next day, primary antibodies were removed by washing three times for 5 min with PBTGS. Cells were stained with secondary antibodies for 1 h in the dark at room temperature and then washed one time for 5 min with each of PBTGS, 0.1 m PB, and 0.05 m PB. The coverslips were then air dried and mounted.
Imaging.
Fluorescence imaging was performed on an AxioImager Z1 microscope (Carl Zeiss) fitted with an apotome and an AxioCam digital camera (Carl Zeiss). AxioVision acquisition software (Carl Zeiss) was used for collection of images. Images in the same dataset were acquired using uniform exposure times. Experiments were performed at least in triplicate. Fluorescence intensity was measured using NIH ImageJ. In some images, contrast and brightness were linearly adjusted using Photoshop (Adobe Systems). Image processing was applied uniformly across all images within a given dataset.
Statistical analyses and quantification.
Two-tailed Student's t tests, with p < 0.05 as significance criteria, were used when appropriate. Results are displayed as mean ± SEM.
Results
Generation and validation of Navβ4-specific antibodies
To determine the subcellular localization of Navβ4, we generated rabbit polyclonal antibodies targeting epitopes within the C-terminal tail of Navβ4 that include the putative open channel blocking peptide sequence (Grieco et al., 2005). In collaboration with the NIH-sponsored monoclonal antibody resource at University of California Davis, NeuroMab, we also generated mouse monoclonal antibodies targeting Navβ4. We designed our antigenic peptides against the C terminus because it is unique among the Na+ channel β subunits and does not share strong sequence identity with other proteins.
To test the specificity of the anti-Navβ4 antibodies, we expressed GFP-tagged Navβ4, V5/His-tagged Navβ1, or V5/His-tagged Navβ2 proteins in COS-1 cells. Navβ1 and Navβ2 share up to 35% sequence identity with Navβ4. Immunostaining of Na+ channel β subunit-transfected COS-1 cells confirmed the specificity of both the mouse monoclonal α-Navβ4 (Fig. 1A) and rabbit polyclonal α-Navβ4 antibodies for Navβ4 (Fig. 1B). Navβ4 immunoreactivity colocalized with GFP in the Navβ4-transfected cultures but was undetectable in the Navβ1–V5/His- and Navβ2–V5/His-transfected cells that showed positive immunoreactivity when immunostained for V5 (Fig. 1A,B) or His (data not shown) tags. Immunoblotting of protein homogenates collected from COS-1 cells transfected with one of the three tagged Navβ subunit constructs (Fig. 1C) showed a single band at ∼63 kDa in the Navβ4–GFP-transfected cell homogenates when immunolabeled with the α-Navβ4 antibodies (Fig. 1C, mouse monoclonal, D, rabbit polyclonal) or an antibody against GFP. To test the specificity of the α-Navβ4 antibodies on neuronal cells, we performed immunoblots on rat whole-brain membrane protein homogenates. By immunoblot, the mouse monoclonal Navβ4 antibody recognized a single band at 38 kDa (Fig. 1D), a molecular mass consistent with previous reports of Navβ4 (Yu et al., 2003). Similarly, the rabbit polyclonal Navβ4 antibody recognized the 38 kDa Navβ4 band. However, this antibody also recognized a nonspecific band around 110 kDa (Fig. 1D, denoted by *). Together, these results demonstrate that the antibodies used in this study recognize Navβ4 and do not cross-react with the Na+ channel β subunits Navβ1 or Navβ2.
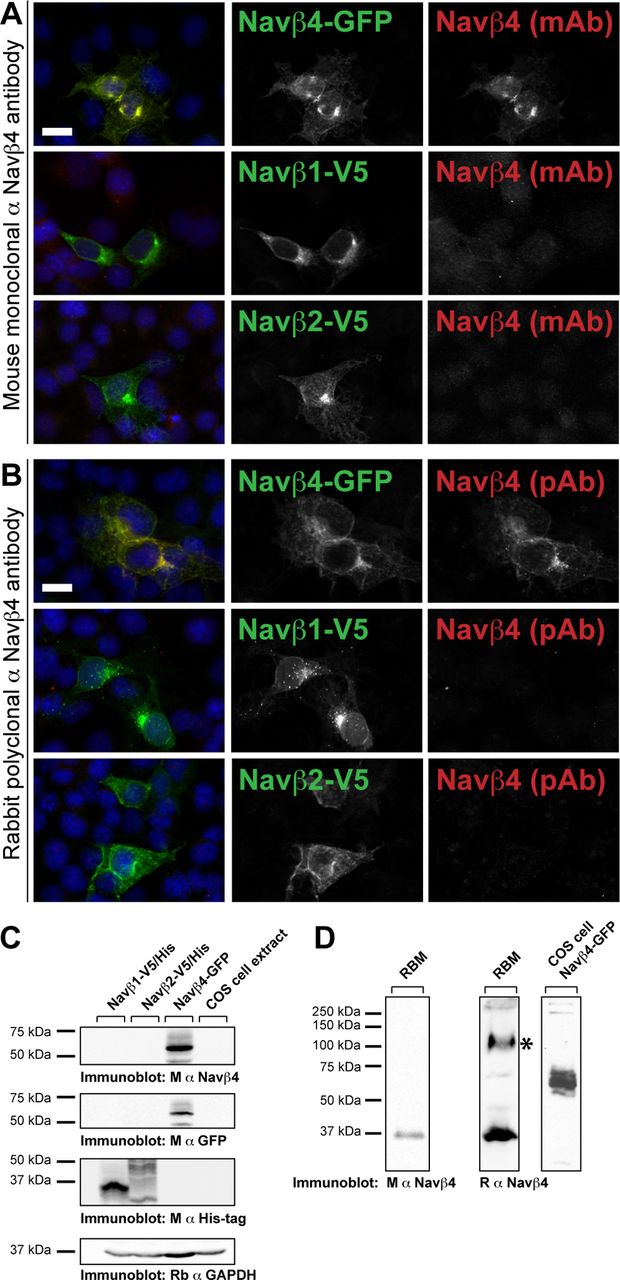
α-Navβ4 antibodies do not cross-react with Navβ1, Navβ2, or other AIS proteins. A, B, COS cells expressing Navβ4–GFP, Navβ1–His/V5, or Navβ2–His/V5 immunostained for either GFP or V5 (green), as appropriate, and Navβ4 (red). A, Mouse monoclonal α-Navβ4. B, Rabbit polyclonal α-Navβ4. Nuclei were labeled with Hoechst stain (blue in the merged image). Scale bars, 20 μm. C, Western blotting of COS cell protein lysates immunoblotted using α-Navβ4 antibodies and antibodies to detect either the GFP- or His-tagged Navβ subunits. D, Immunoblots of rat brain membrane (RBM) protein homogenates or Navβ4–GFP-transfected COS cells, as indicated, probed for Navβ4, which has an approximate molecular weight of 38 kDa. A nonspecific band in the rat brain membrane homogenate recognized by the rabbit polyclonal antibody against Navβ4 is denoted with an asterisk. M, Mouse; R, rat.
Navβ4 is enriched at AIS of diverse cell types
To determine the general expression pattern of Navβ4, we performed Western blots on rat membrane protein homogenates isolated from discrete brain regions. Immunoblotting for Navβ4 revealed single immunoreactive bands at ∼38 kDa in each tissue examined, excluding the optic nerve (Fig. 2A). The lack of Navβ4 detected in the optic nerve likely reflects the low amount of Navβ4 protein relative to the total protein in the homogenate (see also Fig. 4). The wide distribution of Navβ4 protein expression is consistent with previously published reports of Navβ4 expression by RT-PCR and in situ hybridization (Yu et al., 2003).
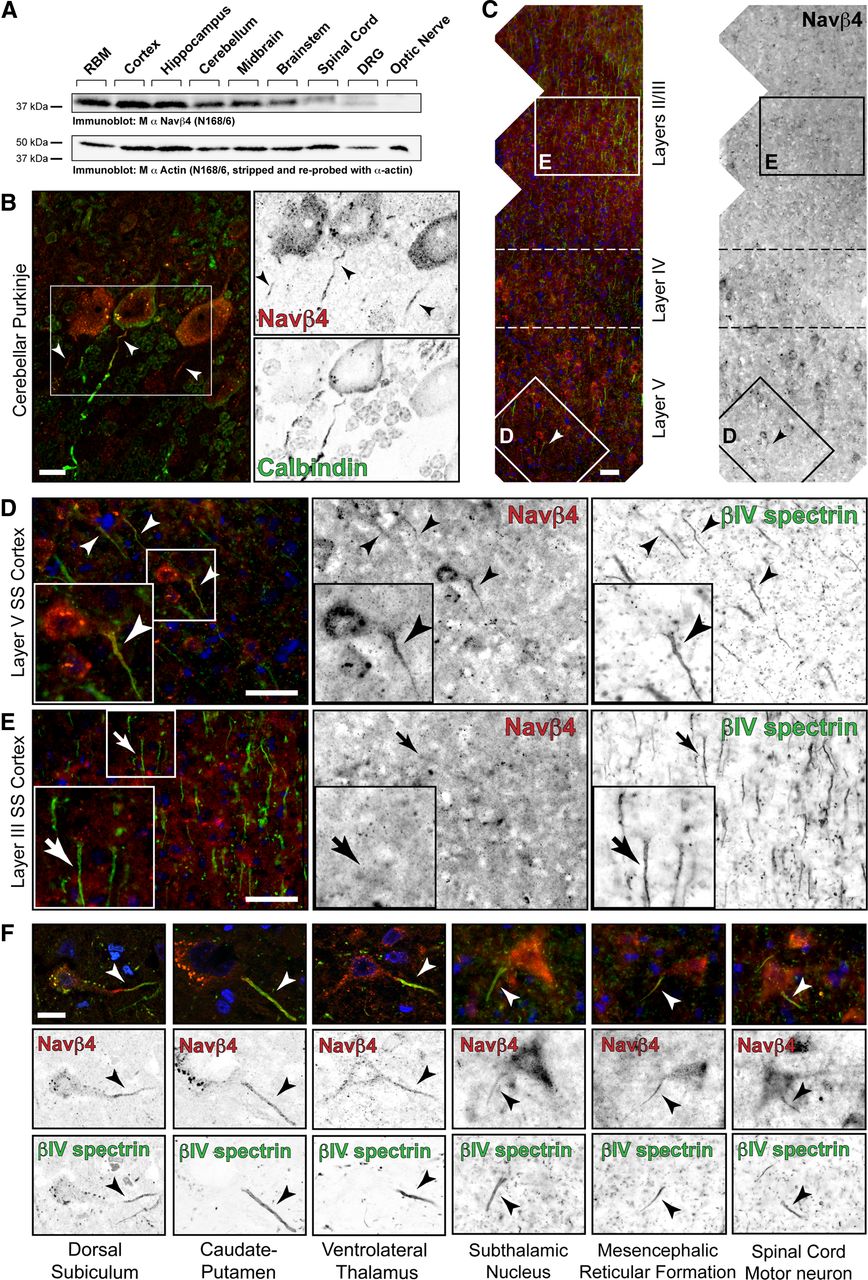
Immunolocalization of Navβ4 reveals AIS enrichment in multiple CNS cell types. A, Western blot of brain-region-specific homogenates probed with the mouse (M) monoclonal α-Navβ4 antibody. Immunoblotting for actin served as a loading control. RBM, Rat brain membrane. B–F, Immunostaining of multiple brain regions for Navβ4 (mouse monoclonal; red) and CB (B, green) or βIV spectrin (C–F, green). Hoechst nuclear staining is shown in blue in the merged images. B, Immunostaining of cerebellar sections for Navβ4 (red) and the Purkinje cell marker CB (green). C, Immunostaining of somatosensory cortex showing layer-specific enrichment of Navβ4 expression. Boxes D and E are expanded in the subsequent panels. F, Representative images of Navβ4-expressing neurons in the indicated brain regions. AIS enrichment is demonstrated by colocalization with βIV spectrin. Scale bars: B, F, 20 μm; C–E, 40 μm.
Resurgent Na+ current has been observed in only a subset of neuron types, and Navβ4 has been proposed to underlie both INaR and INaP (Grieco et al., 2005; Aman et al., 2009). Therefore, we considered whether (1) Navβ4 expression correlates with INaR, (2) Navβ4 expression is most pronounced in cells that fire rapidly, and (3) Navβ4 colocalizes with the dense clusters of sodium channels enriched at sites of AP generation (the AIS) and propagation (nodes of Ranvier). Immunostaining of brain sections revealed markedly heterogeneous cell-type-specific expression of Navβ4 protein at the AIS of many different cell types (Fig. 2B–F). In cerebellar sections, Navβ4 immunoreactivity was evident in 100% of Purkinje neuron somata; of these, 88.3 ± 1.3% (N = 3, n = 100) had enriched Navβ4 immunoreactivity at the AIS (Fig. 2B; Table 1). Interestingly, in the neocortex, Navβ4 was most strongly expressed in layer V pyramidal neurons and neighboring, PV-positive (PV+) interneurons (Figs. 2C–E, 3A, Table 1). Colabeling for the AIS protein βIV spectrin showed that Navβ4 is highly enriched at the AIS of layer V pyramidal neurons (Fig. 2D) but is essentially undetectable at layer II/III neuron AIS (Fig. 2E) (74.0 ± 4.4 vs 5.0 ± 4.4% βIV spectrin+ AIS were Navβ4+, respectively; N = 3, n = 200), suggesting layer-specific mechanisms of Na+ channel modulation. Navβ4 immunoreactivity was also enriched at the AIS of a subset of neurons in the dorsal subiculum, the striatum, the ventrolateral thalamus, the subthalamic nucleus, the mesencephalic reticular formation, and spinal cord motor neurons (Fig. 2F). Thus, although Navβ4 expression is not a fail-safe predictor of INaR (cultured spinal neurons have been reported to express only small, if any, resurgent currents; Pan and Beam, 1999), we identified Navβ4 protein expression in several cell types reported previously to express INaR (Bean, 2007). Notably, Navβ4 immunoreactivity was not detected in hippocampal area CA1 or CA3 pyramidal neuron AIS, which is consistent with the low levels of mRNA expression and the rare observation of INaR in these neuron types (Yu et al., 2003; Castelli et al., 2007a). Together, these results suggest that there is diversity in the molecular composition of the AIS across cell types. To the best of our knowledge, Navβ4 is one of the first modulatory ion channel subunits to be identified as a cell-type-specific AIS protein.
Navβ4 expression in PV/CB-expressing cells

Navβ4 is enriched at interneuron AIS. A–C, Triple immunostaining of Navβ4 (red) (mouse monoclonal antibody) with the calcium binding proteins CB and PV in green or blue, as indicated. Arrowheads highlight Navβ4+ AIS. Scale bars: A, B, 40 μm; C, 80 μm. A, Immunostaining of somatosensory cortex layers V and II/III, as indicated. The Navβ4+ layer V pyramidal neuron AIS is highlighted by the arrow in the top row. The bottom row displays the relatively low expression of Navβ4 among both the excitatory pyramidal neurons and CB and/or PV+ interneurons of layer II/III cortex. B, Immunostaining of Navβ4+ interneurons in the hippocampus. Arrowhead points to the Navβ4+ AIS of the CB/PV+ hippocampal interneuron featured in the top row. The arrowhead in the bottom row highlights the Navβ4+ AIS of the CB-expressing, PV− hippocampal neuron featured in the bottom row. C, Immunostaining of the TB showing Navβ4 expression and AIS-enrichment in the CB/PV-expressing cells, indicated by the arrowheads. Scale bars, 80 μm.
Currently, it remains unknown whether fast-spiking interneurons express Navβ4 protein. Several classes of physiologically distinct interneurons exist throughout the brain. Beyond their anatomical location, these can be further distinguished by their molecular expression profiles and their innervation patterns onto target cell subcellular domains (Klausberger and Somogyi, 2008; Klausberger, 2009). To begin to determine which, if any, interneuron types express Navβ4, we colabeled tissue sections with α-Navβ4 antibodies and antibodies targeting the interneuron-specific calcium-binding proteins PV and CB. Triple immunostaining showed Navβ4 in a reticular pattern surrounding the nucleus as well as AIS-specific enrichment in a subset of cells characterized by PV and/or CB expression (Fig. 3; Table 1). Navβ4 immunoreactivity was evident in 81.6 ± 9.2% of the PV-expressing interneurons in layer V neocortex and at 78.2 ± 2.9 in the hippocampal CA1 region and the subiculum (Figs. 3A,B; Table 1). Layer V PV/CB+ cells uniformly expressed Navβ4 in layer V cortex, but Navβ4 was only enriched at the AIS of 42.9 ± 14.3% of these (Fig. 3A). In layer II/III cortex, Navβ4 was expressed in 40.7% of PV-expressing interneurons but was evident at the AIS of only 6.7 ± 6.7% of these cells [N = 3, n = 52 (N is the number of animals, and n is the number of cells analyzed per animal)] (Fig. 3A). Although PV/CB+ basket cells were infrequently encountered in layer II/III cortex, all of those identified had Navβ4+ AIS. However, PV+ interneurons in the cortex/amygdala transition did not show Navβ4 immunoreactivity (N = 3, n = 10). Among the CB-expressing, PV-negative (PV−) interneurons quantified, those in the hippocampus were the most likely to express Navβ4, with 96.0 ± 1.5% of the population expressing Navβ4, and 70.3 ± 1.9% of those had detectable Navβ4 immunoreactivity at the AIS (N = 3, n = 52) (Fig. 3B; Table 1). CB+ neurons of the inferior colliculus and other brainstem regions had strong somatic and AIS-enriched Navβ4 immunoreactivity, consistent with the findings of Kodama et al. (2012). Our data suggest that a subset of fast-spiking interneurons, including PV/CB+ interneurons of the cortex and hippocampus, express Navβ4 and that many of these have AIS enrichment of Navβ4; however, we also observed Navβ4 expression in CB-expressing, PV− cells that have been reported to fire at uniformly low rates (Hartwich et al., 2009). Quantification of Navβ4 expression in interneuron populations of discrete brain regions is provided in Table 1.
During these studies, we also noted strong expression of Navβ4 in the somata and at the AIS of the PV/CB-expressing neurons of the trapezoid body (TB) [99.0 ± 1.0% PV+/CB+ TB neurons were Navβ4+, whereas Navβ4 was specifically enriched at the AIS of 66.7 ± 0.6% of these (N = 3, n = 52)] (Fig. 3C,D). Therefore, Navβ4 may contribute to the resurgent sodium currents displayed by principle neurons in the medial nucleus of the TB, which are thought to be modulated by factors other than α subunits (Leão et al., 2006). Together, these data highlight the heterogeneity of Navβ4 expression among cell types.
Navβ4 is enriched at nodes of Ranvier
In myelinated axons, APs are regenerated at nodes of Ranvier. Nodes and the AIS share a common molecular composition, including dense clusters of voltage-gated Na+ channels that underlie the depolarizing phase of the AP. Furthermore, because some peripheral sensory neurons can support trains of APs at frequencies approaching 1 kHz (Wellnitz et al., 2010), we wondered whether Navβ4 also modulates Na+ channels enriched at nodes. To test this idea, we immunostained CNS and PNS nerves with antibodies against Navβ4, the nodal marker βIV spectrin, and juxtaparanodal Kv1.2 channels (Fig. 4A). We found that Navβ4 immunoreactivity colocalized with βIV spectrin at a subset of optic nerve (31.3 ± 2.7%), ventral root (88.3 ± 1.9%), and dorsal root (72.3 ± 3.8%) nodes of Ranvier (Fig. 4B). On closer examination, we determined that there is a small but statistically significant increase in the probability of Navβ4+ immunoreactivity at nodes of large- versus small-diameter axons (82.0 ± 1.0 vs 67.5 ± 4.2% Navβ4+, respectively; p = 0.015) (Fig. 4C,D). Axons ≥4 μm in diameter were counted among the large axons, whereas those ≤3 μm in diameter were classified as small. No significant difference was found between the probabilities of Navβ4 enrichment at large versus small ventral root axons (90.7 ± 1.5 vs 92.3 ± 2.3%, respectively; p = 0.58) (Fig. 4C,D). These results demonstrate that Navβ4 is enriched at a subset of nodes of Ranvier in both the CNS and PNS and suggest that Navβ4 modulation of nodal Na+ channels may contribute to either INaP or INaR at nodes of Ranvier during AP propagation.

Navβ4 is enriched at a subset of CNS and PNS nodes of Ranvier. A, Immunostaining of rat dorsal and ventral spinal cord roots and optic nerve using antibodies against Navβ4 (rabbit polyclonal; green), nodal βIV spectrin (blue), and juxtaparanodal Kv1.2 channels (red). B, Quantification of the percentage of nodes with detectable Navβ4 immunoreactivity enrichment in the indicated nerve types (n = 100, N = 3). C, Quantification of Navβ4+ nodes in large- vs small-diameter ventral (VR) and dorsal (DR) root axons (n = 100, N = 4). *p < 0.05, ***p < 0.001. Error bars represent SEM. D, Images show representative examples of Navβ4+ and Navβ4− large- and small-diameter dorsal root nodes coimmunostained for βIV spectrin (blue) and Kv1.2 (red), shown in the merged images. Scale bars, 10 μm.
AIS localization of Navβ4 requires ankG and Na+ channel α subunit expression
Na+ channel α subunit recruitment to AIS and nodes of Ranvier is mediated through an ankyrin-binding motif located within the II–III cytoplasmic linker of the channels (Garrido et al., 2003; Lemaillet et al., 2003; Gasser et al., 2012). Similarly, neurofascin-186 and βIV spectrin localization to the AIS also require binding to ankG (Zhang et al., 1998; Yang et al., 2007). To determine how Navβ4 is recruited to the AIS, we first determined whether FL GFP-tagged Navβ4 constructs localize to the AIS in cultured hippocampal neurons. Cells were transfected at the time of plating and allowed to mature DIV14 before analysis. Immunostaining of transfected neurons revealed AIS-specific enrichment of Navβ4–GFP protein (Fig. 5A). In contrast, and consistent with our immunofluorescence results from brain sections, we did not detect any endogenous Navβ4 expression in cultured hippocampal pyramidal neurons (Fig. 5B). Thus, exogenously introduced GFP-tagged Navβ4 protein can localize to the AIS without additional factors unique to INaR-expressing cells.

AnkG and Nav1 α subunit expression are required for Navβ4 AIS targeting. A, Hippocampal neurons transfected with GFP-tagged Navβ4 and immunostained for GFP (green), ankG (red), and MAP2 (blue). B, Cultured hippocampal neurons immunolabeled for Navβ4 (green), ankG (red), and MAP2 (blue). C, D, Immunostaining of cultured hippocampal neurons cotransfected with the indicated shRNA and FL Navβ4–GFP constructs. Neurons were labeled with α-βIV spectrin or α-neurofascin-186 (blue), α-GFP (green), and α-ankG, or α-pan Nav channels (red). Transfected cells are marked with an *. Scale bars, 20 μm. C, Immunostaining of cultured neurons transfected with shRNA targeting the AIS-organizing protein ankG or Na+ channel α subunits, as indicated. Arrowhead indicates βIV spectrin+/Navα−/Navβ4− AIS in the bottom row, and arrows point toward neighboring control neuron AIS in all panels. D, Knockdown of the AIS proteins βIV spectrin and neurofascin-186. Arrowheads highlight the location of Navβ4+ AIS in all panels.
To determine whether the subcellular distribution of Navβ4 depends on ankG, we cotransfected cells with FL GFP–Navβ4 and either ankG-targeted or control shRNA constructs. Knockdown of ankG disrupted Navβ4 targeting to the AIS in 100% of transfected neurons (N = 3, n = 100; Fig. 5C). To test whether the ankG-dependent recruitment of Na+ channel α subunits is required for Navβ4 enrichment at the AIS, we cotransfected GFP-tagged Navβ4 with Nav1-targeted shRNA. Navβ4 AIS localization was completely abolished in the absence of Nav1 α subunit expression, indicating that this interaction is required for Navβ4 recruitment to the AIS (Fig. 5C). Thus, although ankG expression is necessary for AIS localization of Navβ4, it is not sufficient for it but instead requires coexpression and AIS localization of Na+ channel α subunits. However, knockdown of other AIS proteins, including βIV spectrin and neurofascin-186, had no effect on the subcellular distribution of Navβ4 (Fig. 5D). These data show that Navβ4 enrichment at the AIS requires Na+ channel α subunits.
Cysteine 28 is essential for Navβ4 recruitment to the AIS
Similar to Navβ1 and Navβ2, Navβ4 comprises an extracellular Ig-like fold, a single transmembrane domain, and a short C-terminal tail. The N-terminal Ig-like fold of β4 is structurally similar to those of β1 and β2. The extracellular domains of the β subunits are thought to be critical to their function; indeed, β1 has been proposed to function not only in Na+ channel modulation but also in cell adhesion with a role in neurite outgrowth during postnatal CNS development (Brackenbury et al., 2008). The C-terminal tail of Navβ4 is unique among the Navβ subunits and contains the putative open channel blocking peptide.
To determine how Navβ4 interacts with Na+ channel α subunits, we expressed FL and mutant GFP-tagged Navβ4 constructs in cultured hippocampal neurons. First, we transfected terminal truncation mutants in which either the C or N terminus of Navβ4 was deleted. Like FL Navβ4–GFP, we found Navβ4ΔC–GFP enriched at the AIS of 100% of transfected neurons (Fig. 6A). In contrast, the N-terminal deletion completely abolished the AIS localization of Navβ4 (Fig. 6A). Therefore, the determinants of AIS targeting in Navβ4 are contained within its extracellular N-terminal domain.

Cysteine 28 is required for Navβ4 AIS localization. A, Neurons transfected with FL, ΔC, or ΔN Navβ4 constructs and immunostained for GFP (green), ankG (red), and MAP2 (blue). Longitudinal line scans through the AIS show fluorescence intensity (F.I.) of the Navβ4–GFP (green), ankG (red), and MAP2 (blue) signals. B–E, Immunostaining of cultured hippocampal neurons transfected with Navβ4-C28A (B) or the indicated Navβ4 single point mutants (C) (C23A, C101A, and G52A). Neurons were immunolabeled using antibodies against GFP (green), ankG (red), and MAP2 (blue). Scale bar, 20 μm. D, Quantitative analysis of Navβ4 point mutant targeting to the AIS in cultured hippocampal neurons. E, Western blotting for Navβ4 in whole-brain membrane protein homogenates denatured in reducing (R) or non-reducing (NR) sample buffer. TM, Transmembrane.
The N terminus of Navβ4 has three cysteine residues. By analogy to β1 and β2, two of these, C23 and C101, are thought to provide an internal disulfide bond to stabilize the quaternary structure of the extracellular domain (Barbieri et al., 2012). However, the third cysteine, C28, is left unpaired with the potential to form an extra-molecular covalent disulfide bond (Yu et al., 2003). Single point mutation of C28 to alanine (Navβ4–C28A) resulted in disrupted Navβ4 targeting, similar to the ΔN results (Fig. 6B,D). For additional analysis, we generated single point mutations of C23 and C101 as well as amino acids that were either within close proximity or well separated from the cysteines. Mutation of either paired cysteine, C23 or C101, also disrupted AIS localization of Navβ4 (Fig. 6C,D), presumably as a result of disruption of the Ig-like fold causing C28 to no longer be positioned for favorable interaction with the reciprocal α subunit cysteine. Single point mutation of G30 and L21 to alanine also caused mislocalization of Navβ4, whereas mutation of Navβ4 at G52 (Fig. 6C), T24, T45, V63, and V103 preserved AIS localization (Fig. 6D).
In support of the idea that Navβ4 and Nav1 α subunits form a covalent disulfide linkage, we found that a single 38 kDa immunoreactive band was present in Western blots of brain membrane homogenates denatured in reducing sample buffer and probed for Navβ4. In contrast, when the proteins were denatured in non-reducing sample buffer, two bands were evident after probing for Navβ4: the first at 38 kDa, representing Navβ4 alone, and the second at a molecular weight (>250 kDa) corresponding to Navβ4 linked to an α subunit (Fig. 6E). These findings are consistent with previous reports (Yu et al., 2003). Together, the data presented in Figures 5 and 6 demonstrate that Navβ4 recruitment to the AIS depends on its direct interaction with Nav1 α subunits through a disulfide bond mediated by C28 in Navβ4.
Navβ4 localization at nodes of Ranvier requires disulfide linkage to α subunits
Although they share a common molecular organization, recent studies show that the mechanisms governing AIS assembly differ from those at nodes (Hedstrom et al., 2008; Feinberg et al., 2010; Zhang et al., 2012). ankG localization in the proximal axon is essential for AIS formation because all other AIS proteins, including Na+ channels, are recruited to the AIS through either direct or indirect linkage to ankG (Hedstrom et al., 2008). Whereas ankG clustering at the AIS is specified completely by the neurons, Na+ channel clustering at nodes of myelinated axons depends on neuron–glia interactions (Eshed et al., 2005; Feinberg et al., 2010; Galiano et al., 2012; Zhang et al., 2012). It is possible that Navβ4 localization to nodes occurs independently of Na+ channel localization, because previous reports suggest that Navβ1 directly interacts with other axonal and glial cell adhesion molecules, including neurofascin-186 and NrCAM (McEwen and Isom, 2004). To determine whether a disulfide-mediated interaction between Navβ4 and Na+ channel α subunits is necessary for Navβ4 clustering along myelinated axons, we used an in vitro DRG neuron–Schwann cell myelinating coculture system. Purified DRG neurons were prepared from embryonic rat pups in parallel with purified Schwann cells from P2 rats. DRG neurons were transfected with either the FL, ΔC, ΔN, or C28A GFP-tagged Navβ4 expression constructs on DIV10 and then combined with Schwann cells at DIV11. Myelination was induced 1 week later. After 2 weeks of myelination, we fixed the neurons and immunostained for GFP, the myelin marker MBP, and nodal marker ankG. Similar to our AIS results, we found that the FL and ΔC Navβ4–GFP proteins colocalized with ankG at heminodes (heminodes are precursors to nodes of Ranvier) (heminodes FL Navβ4+ = 94.5 ± 3.9%, n = 66, N = 3; heminodes Navβ4ΔC+ = 94.6 ± 2.6%, n = 76, N = 3) (Fig. 7A,C). In contrast, we found that the ΔN- and C28A–Navβ4–GFP proteins were uniformly distributed throughout the axon and failed to become enriched at ankG+ heminodes (heminodes Navβ4ΔN+ = 8.1 ± 3.6%, n = 138, N = 3; heminodes Navβ4C28A+ = 9.1 ± 5.9%, n = 42, N = 3) (Fig. 7B,C). These results show that the recruitment of Navβ4 to nodes of Ranvier, as with the AIS, depends on its direct interaction with Na+ channel α subunits through a disulfide bond at C28.

Cysteine 28 is required for Navβ4 targeting to nodes of Ranvier. DRG neurons transfected with Navβ4 expression constructs encoding FL-, ΔC- (A), ΔN-, and C28A–Navβ4–GFP (B) constructs cocultured with myelinating Schwann cells. Myelinated axon segments were detected by immunostaining for MBP (red), whereas ankG immunoreactivity (blue) defined heminodes and forming nodes of Ranvier. The FL or mutant GFP-tagged Navβ4 proteins were detected by GFP immunoreactivity (green). Scale bar, 10 μm. C, Quantification of FL-, ΔC-, ΔN-, and C28A–Navβ4+ heminodes in the myelinating cocultures.
Discussion
Open channel block by the C-terminal tail of Navβ4 has been proposed as a potential mechanism for the generation of INaR in neurons (Grieco et al., 2005; Bant and Raman, 2010). Na+ currents with resurgent kinetics have been suggested to stimulate neuronal excitability and promote repetitive firing (Khaliq et al., 2003). Extracellular recordings revealed that APs are first initiated in the AIS during both single-spike and bursting activity (Kole, 2011); hence, local regulation of Na+ channel activity at the AIS is central to controlling neuronal firing properties. Here, we investigated the neuronal expression pattern and subcellular localization of Navβ4. Our results link Navβ4 expression and subcellular localization at the AIS and nodes of Ranvier to its proposed role in the generation of INaR and INaP and, consequently, to its contribution to repetitive spiking in neurons.
Patch-clamp experiments using focal application of TTX in perirhinal cortex suggest that INaR is primarily generated in the AIS (Castelli et al., 2007b). Notably, quantitative freeze-fracture immunogold studies of Nav1.6, the Na+ channel α subunit reported to carry the bulk of INaR in some cells (Raman et al., 1997; Khaliq et al., 2003; Grieco and Raman, 2004), suggest a 40-fold enrichment of Nav1.6 at the AIS over the somatodendritic domain (Lorincz and Nusser, 2010). Thus, we reasoned that, if Navβ4 is necessary for the generation of INaR in multiple cell types, then it should be strategically positioned to modulate the Na+ channels involved in AP initiation, namely those enriched at the AIS. Using specific antibodies recognizing a unique epitope in the C-terminal tail of Navβ4 (Fig. 1), we found cell-type-specific enrichment of Navβ4 at the AIS of neuron classes known to express INaR, including cerebellar Purkinje neurons and subthalamic neurons, among others (Fig. 2). Furthermore, we observed layer-specific enrichment of Navβ4 at the AIS of cortical pyramidal neurons in layer V somatosensory cortex that was mostly absent from AIS in layers II/III, IV, and VI cortical pyramidal neurons. We also found Navβ4 expression at the AIS of fast-spiking, PV+ cortical interneurons and CB+ brainstem interneurons (Fig. 3); however, Navβ4 was also expressed in a subset of CB-expressing, PV− neurons, suggesting that its expression is not restricted to high-frequency firing cell types. Navβ4 protein expression in brainstem interneurons is consistent with recent single-cell transcript profiling results that found Navβ4 expression to be a key marker differentiating fast-spiking inhibitory neurons from neighboring cells in the medial vestibular nucleus (Kodama et al., 2012). Finally, we observed strong Navβ4 immunoreactivity in the TB, a site in which developmental increases in INaR, perhaps facilitated by Navβ4 expression, contributes to rapid signal generation in these cells (Leão et al., 2006). Thus, Navβ4 is enriched at the site of AP initiation (the AIS) of cell types characterized by high-frequency AP generation, some of which are known to carry INaR.
Similar to the AIS, Na+ channels are densely clustered at nodes of Ranvier in myelinated axons for the efficient propagation of the AP. Here, we found that Navβ4 is enriched at a subset of CNS and PNS nodes (Fig. 4). To our knowledge, INaR has not been directly recorded from nodes; however, non-inactivating channel kinetics underlying INaP have been measured at frog nodes of Ranvier (Dubois and Bergman, 1975), and some sensory neuron nodes in the DRG are capable of faithfully propagating AP trains approaching 1 kHz over long distances that can exceed 1 m (Wellnitz et al., 2010). We propose that the molecular mechanisms underlying fast spike generation at AIS are likely conserved at nodes. Intriguingly, a recent report on the contribution of nodes of Ranvier to repetitive firing revealed that nodal Na+ channel activity critically contributes to burst probability in neocortical pyramidal neurons (Kole, 2011). This study showed that activity at the first node of Ranvier is essential for burst generation and that subthreshold INaP from distal axonal nodes of Ranvier contributes to the overall level of neuronal excitability. Whether or not INaR is expressed at nodes of Ranvier and the specific contribution of Navβ4 to AP propagation at nodes remain active areas of investigation.
The contribution of Navβ4 to repetitive firing may not be restricted to its role at the AIS nor even specifically in INaR but may also extend to facilitating INaP generation at nodes of Ranvier distributed throughout the entire length of the axon. Furthermore, it is possible that Navβ4-modulated Na+ channel gating at nodes of Ranvier may partially account for the molecular basis of subthreshold axonodal INaP generation. Facilitating INaP may be an additional mechanism by which Navβ4 contributes to repetitive firing in neurons. Finally, resurgent current and Navβ4 expression alone may not guarantee repetitive high-frequency firing. Instead, high-frequency firing likely depends not only on resurgent Na+ current but also on the complement of high-threshold, rapidly deactivating K+ currents found in specific neuron types.
By studying Navβ4 trafficking to the AIS of cultured hippocampal pyramidal neurons that do not normally express Navβ4, we were able to show that it can be recruited to the AIS without any special component found only in Navβ4-expressing neurons. Specifically, our results show that Navβ4 subunits are recruited to the AIS and nodes of Ranvier through a direct covalent disulfide bond with voltage-gated Na+ channel α subunits, similar to Navβ2 (Catterall, 1999; Yu et al., 2003). Although it remains unknown whether Navβ4 binds to the α subunits before trafficking to these domains, our data suggest that any additional molecular interactions Navβ4 may have with nodal or AIS cell adhesion molecules or ECM proteins are insufficient to either recruit or stabilize Navβ4 at these sites. It will be interesting to determine which extracellular cysteine on the Na+ channel α subunit is responsible for the interaction with Navβ4. Although site-directed mutagenesis of FL Na+ channels is technically quite difficult, it is possible and has been used to study Na+ channel localization in vivo (Gasser et al., 2012). However, sequence alignment of brain Na+ channels reveals 12 conserved extracellular cysteines, making it difficult to predict which of these forms the disulfide bond with Navβ4. Furthermore, among the known variants of Na+ channels, none affect an extracellular cysteine. Thus, determining where Navβ4 interacts with the Na+ channel α subunit will require future analyses of many Na+ channel mutants.
Navβ4 was discovered by mining expressed sequence tag databases for novel sequences with high homology to Navβ2 (Yu et al., 2003). Examination of the human genome reveals that the family of Na+ channel β subunit isoforms likely arose from a series of gene duplication events resulting in the four distinct transcripts. Given the heterogeneous expression pattern of Navβ4, it will be interesting to uncover the mechanisms regulating transcriptional control of the various Navβ subunits. A limited phylogenetic analysis across 13 vertebrate species revealed conservation of key amino acid residues in the blocking peptide sequence of the C terminus of Navβ4 (Lewis and Raman, 2011), suggesting that repetitive firing in Purkinje neurons, facilitated by Navβ4-mediated resurgent kinetics, was selected for in vertebrates. Together, our data suggest that neuronal firing pattern diversity arises not only from variability in the expression of ion channel α subunits but also in cell-type-specific regulation of auxiliary protein expression. Furthermore, the specific subcellular localization of modulatory ion channel subunits, e.g., Navβ4 targeting to AIS and nodes, likely determines the functional impact they have on neuronal computation (Nusser, 2009).
In summary, the data presented here identify Navβ4 as a cell-type-specific component of the AIS and nodes of Ranvier and suggest a correlation between Navβ4, resurgent Na+ current, and repetitive firing. We also determined that Navβ4 is recruited to these axonal electrogenic domains through direct interaction with the voltage-gated Na+ channels enriched here. Our findings contribute to the idea that variability in the cell-type-specific expression pattern and subcellular distribution of ion channels and their modulatory subunits increases neuronal diversity, a key component underlying the evolution of higher cognition (Lorincz and Nusser, 2008; Nusser, 2009; Debanne et al., 2011).
Footnotes
This work was supported by National Institutes of Health Grants NS073295 (S.A.B.) and NS044916 (M.N.R.). We thank Dr. James Trimmer and NeuroMab for generating the monoclonal α-Navβ4 antibody and Dr. Yasuhiro Ogawa for generating the FL GFP-tagged Navβ4 construct. We also thank Dr. Indira Raman and Dr. Maarten Kole for helpful discussions.
- Correspondence should be addressed to Dr. Matthew N. Rasband, Department of Neuroscience, Baylor College of Medicine, One Baylor Plaza, BCM295, Houston, TX 77030. rasband{at}bcm.edu




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}