

Abstract
Cytoplasmic Ca2+ elevation and changes in Rho GTPase activity are both known to mediate axon guidance by extracellular factors, but the causal relationship between these two events has been unclear. Here we show that direct elevation of cytoplasmic Ca2+ by extracellular application of a low concentration of ryanodine, which activated Ca2+ release from intracellular stores, upregulated Cdc42/Rac, but downregulated RhoA, in cultured cerebellar granule cells and human embryonic kidney 293T cells. Chemoattractive turning of the growth cone triggered by a gradient of ryanodine was blocked by overexpression of mutant forms of Cdc42 but not of RhoA in Xenopus spinal cord neurons. Furthermore, Ca2+-induced GTPase activity correlated with activation of protein kinase C and required a basal activity of Ca2+/calmodulin-dependent protein kinase II. Thus, Rho GTPases may mediate axon guidance by linking upstream Ca2+ signals triggered by guidance factors to downstream cytoskeletal rearrangements.
Introduction
Pathfinding of growing axons in the developing nervous system is guided by extracellular factors, which trigger a cascade of cytoplasmic signaling events that determine the direction of growth cone extension. As a second messenger in the cytoplasm, Ca2+ plays critical roles in regulating motility of the axonal growth cone (Kater et al., 1994; Gomez et al., 2001) and in the guidance of growth cone extension by many extracellular factors, including netrin-1, brain-derived neurotrophic factor (BDNF), myelin-associated glycoprotein (MAG), and N-cadherin (Hong et al., 2000; Ming et al., 2001; Chadborn et al., 2002; Wong et al., 2002; Henley and Poo, 2004). A gradient of intracellular Ca2+ concentration ([Ca2+]i) created either by exposure to an extracellular gradient of ryanodine, which activates Ca2+ release from cytoplasmic stores (Hong et al., 2000), or by local photo-activated Ca2+ release from a caged Ca2+ compound (Zheng, 2000) is sufficient to trigger growth cone turning, suggesting that Ca2+ indeed can mediate the guidance signal.
Growth cone turning requires rearrangement of cytoskeleton at the growth cone. The Rho family of small GTPases may serve as “molecular switches” to regulate the cytoskeletal rearrangements involved in many forms of cell motility (Etienne-Manneville and Hall, 2002). Many factors that influence axon growth, including slits, semaphorins, ephrins, netrins, neurotrophins, MAG, and Nogo, are known to regulate intracellular activity of Rho GTPases (Wahl et al., 2000; Whitford and Ghosh, 2001; Wong et al., 2001; Li et al., 2002; Wang et al., 2002; Yuan et al., 2003), and the perturbation of the activity of Rho GTPases in vivo (Dickson, 2001) or in vitro (Yuan et al., 2003) leads to axon pathfinding defects. Because Cdc42/Rac positively regulate axon extension and stabilization, whereas RhoA increases the tendency of growth cone retraction, it is likely that an activity gradient of Cdc42/Rac across the growth cone triggered by extracellular guidance cues results in preferential axon extension toward the side of higher Cdc42/Rac activity, whereas higher RhoA activity results in preferential retraction of filopodia and repulsive growth cone turning. In this model, the activity of Rho GTPases is not merely permissive for growth cone guidance, but directly mediates the guidance signal.
Previous studies have shown that elevation of [Ca2+]i regulates the activities of Rho GTPases in various cell types (Fleming et al., 1998, 1999; Price et al., 2003). However, Rho GTPase activities have also been shown to regulate the [Ca2+]i dynamics (Wilk-Blaszczak et al., 1997; Costello et al., 1999; Djouder et al., 2000; Singleton and Bourguignon, 2002). In view of the essential role of both Ca2+ and Rho GTPases in growth cone guidance, it is important to determine their causal relationship in transducing guidance signal. Using ryanodine to directly trigger intracellular [Ca2+]i elevation, we showed in the present work that Rho GTPases can function downstream of Ca2+ signals to mediate growth cone turning, although GTPases activity can also affect slightly the Ca2+ signal. Furthermore, we found that the Ca2+ effector enzyme protein kinase C (PKC) is essential for the Ca2+-dependent regulation of Rho GTPases during the transduction of guidance signals.
Materials and Methods
Growth cone turning assay. Cultures of Xenopus spinal neurons were prepared from 1-d-old Xenopus embryos by methods described previously (Wang et al., 1998). Briefly, culture medium consisted of 50% (v/v) Leibovitz L-15 medium (Invitrogen, Gaithersburg, MD), 1% (v/v) fetal bovine serum (HyClone, Logan, UT), and 49% (v/v) Ringer's solution (in mm: 115 NaCl, 2 CaCl2, 2.5 KCl, and 10 HEPES, pH 7.4). The cells were used for experiments 14 h after plating at room temperature (20-22°C). To assay growth cone turning, the pipette tip (1 μm opening) was placed 100 μm away from the center of the growth cone of an isolated neuron and at an angle of 45° with respect to the initial direction of neurites (indicated by the last 10 μm segment of the neurite). A standard pressure pulse of 3 psi was applied at a frequency of 2 Hz with durations of 20 ms. Theoretical analysis (Lohof et al., 1992) and direct measurements of the gradient using fluorescent dyes (Zheng et al., 1994) have shown that, using standard methods, the average concentration of the chemical is ∼103-fold lower at the growth cone than that in the pipette. The turning angle was defined by the angle between the original direction of neurite extension and a straight line connecting the positions of the growth cone at the onset and the end of the 1 h period. Neurite extension was quantified by measuring the entire trajectory of net neurite growth over the 1 h period. Microscopic images of neurites were captured with a CCD camera (JVC TK-1381; Victor Company, Yokohama, Japan) attached to a phase-contrast microscope (CK40; Olympus Optical, Tokyo, Japan) and stored in a computer for later analysis using Scion (Frederick, MD) Image programs. To determine the total length of neurite extension, the entire trajectory of the neurite at the end of the 1 h period was measured with a digitizer. Only those growth cones with net extension >5 μm over the 1 h period were included for analysis of turning angles. All turning experiments were performed at room temperature in modified Ringer's solution (in mm: 140 NaCl, 2.5 KCl, 1 MgCl2, 1 CaCl2, and 10 HEPES, pH 7.40). For pharmacological pretreatments, chemicals were added to the culture medium at 30 min before and were present throughout the experiments, unless indicated otherwise. Data are presented as mean ± SEM. Statistical significance was analyzed by Kolmogorov-Smirnov test.
Ca2+ imaging. For Ca2+ imaging, cells were rinsed before being loaded with 2 μm fluo-4 AM and 2 μm fura-Red AM (Molecular Probes, Eugene, OR) with 0.1% dimethylsulfoxide (DMSO) in modified Ringer's solution (MR) for 30 min at room temperature, and then incubated for another 30 min in MR after rinsing twice in MR. Imaging was performed on a Zeiss (Oberkochen, Germany) LSM 510 confocal imaging system with a 40× oil object (Plan-Neofluar, 1.3 numerical aperture; Zeiss). Excitation was provided by the 488 nm line of the laser, and the intensity of the fluorescence between 505 and 550 nm was measured as fluo-4 signal, whereas longer than 635 nm was simultaneously detected as fura-Red signal. Images were acquired with 15 s intervals for frame scans or with 10 ms intervals for line scans. The Ca2+ signals were measured by using the change in ratio of fluo-4/fura-Red relative to the baseline before applying ryanodine (ΔR/R0) in selected regions. To measure the Ca2+ in growth cone under a gradient of specific reagents, the gradient was applied using the standard protocol for turning assay. In some experiments, cells were incubated in BAPTA-AM (10 μm; Calbiochem, La Jolla, CA) or thapsigargin (10 μm; Calbiochem) for at least 30 min, or in toxin B (5 ng/ml; Calbiochem) overnight before measurement.
Western blotting. Different cultures in 35 mm dishes were lysed in 0.2 ml lysis buffer (0.1% SDS, 1% NP-40, 50 mm HEPES, pH 7.4, 2 mm EDTA, 100 mm NaCl, 5 mm sodium orthovanadate, 40 μm p-nitrophenyl phosphate, and 1% protease inhibitor mixture set I; Calbiochem). The lysates were centrifuged at 12,000 rpm for 25 min. The supernatant was collected and denatured. We loaded 20 μg of total protein in each lane, separated in 15% SDS-PAGE and blotted onto polyvinylidene difluoride membrane. The blot was blocked for 3 h at room temperature in 0.2% block-1 (Tropix, Foster City, CA), followed by incubation overnight at 4°C in rabbit polyclonal antibodies [p-PKCα/β (Cell Signaling Technology, Beverly, MA); p-Ca2+/calmodulin-dependent protein kinase II (p-CaMKII; Upstate Group, Waltham, MA)]. Membranes were rinsed and incubated for 1 h with peroxidase-conjugated goat anti-rabbit IgG (1:10,000; Bio-Rad, Hercules, CA). Chemiluminent detection was performed with the ECL kit (Pierce, Rockford, IL). To show equal loading of each lane, the membranes were washed in stripping buffer (2% SDS, 62.5 mm Tris-HCl, pH 6.8, and 100 mm 2-mercaptoethanol) at 70°C for 30 min, rinsed, and reprobed with mouse monoclonal antibodies against actin (Chemicon, Temecula, CA).
Assay for GTPase activity. We measured GTPase activity by methods as described previously (Ren et al., 1999). Briefly, lysates of human embryonic kidney 293T (HEK293T) cells transfected with wild-type Rho GTPases, cultured cerebellar granule cells (postnatal day 7), or cortical neurons (embryonic day 15) were preincubated with glutathione-Sepharose 4B beads (Amersham Biosciences, Buckinghamshire, UK) at 4°C for 1 h with constant rocking and then centrifuged to remove the nonspecific binding. The supernatants were incubated with the bacterially produced glutathione S-transferase (GST)-fused Rac/Cdc42-binding domain of Pak (GST-PBD; 5 μg) or the Rho-binding domain of Rhotekin (GST-RBD; 20 μg) bound to glutathione-coupled Sepharose beads at 4°C for 1 h with constant rocking. The beads and proteins bound to the fusion protein were washed five times with lysis buffer at 4°C, eluted in SDS sample buffer, and analyzed for bound Cdc42, Rac1, or RhoA by Western blotting using mouse monoclonal antibodies against Cdc42 (Santa Cruz Biotechnology, Santa Cruz, CA), mouse monoclonal antibodies against Rac1 (Upstate Group) or mouse monoclonal antibodies against RhoA (Santa Cruz Biotechnology). Protein samples of one-tenth of total lysates were shown to indicate similar loading by Western blotting. For pharmacological pretreatments, chemicals were added to the culture medium 30 min before and were present throughout the experiments. The relative levels of Rho GTPase activation were measured by densitometry analysis of the blots.
Expression of Rho-family GTPases by cDNA blastomere injection. Green fluorescent protein (GFP)-fusion constructs of constitutively active (CA) V12 Cdc42, and dominant-negative (DN) N17 Cdc42, N19 RhoA are gifts from Dr. Gary Bokoch (Scripps Research Institute, La Jolla, CA). Endofree Plasmid Maxi kit (Qiagen, Hilden, Germany) was used to purify the cDNA clones. The final concentration of cDNAs for injection is 0.2 μg/μl for DN-Cdc42 or CA-Cdc42 and 0.5 μg/μl for DN-RhoA and GFP. Approximately 1.5 ng of DN-Cdc42 or CA-Cdc42 and 4 ng of DN-RhoA or enhanced GFP were injected into one of the blastomeres of Xenopus embryos at the two-cell or four-cell stage using an Eppendorf (Hamburg, Germany) transjector 5246, as described previously (Yuan et al., 2003). The embryos were kept in 10% Ringer's solution at room temperature (20-22°C) for 24 h before preparation of the culture. Embryos with GFP fluorescence in spinal cord were selected under the fluorescence microscope for the neuronal culture.
Source and preparation of reagents. Antibodies were purchased from the following sources: rabbit polyclonal antibody against p-PKCα/β from Cell Signaling Technology, rabbit polyclonal antibody against p-CaMKII from Promega (Madison, WI), mouse monoclonal antibody against Cdc42 and mouse monoclonal antibody against RhoA from Santa Cruz Biotechnology, mouse monoclonal antibody against Rac1 from Upstate Group, and mouse monoclonal antibody against actin from Chemicon. BDNF and netrin-1 were purchased from R&D Systems. Ryanodine was from Alomone Laboratories (Jerusalem, Israel), and fluo-4 AM and fura-Red AM were from Molecular Probes (Eugene, OR). All other chemicals were purchased from Calbiochem. Netrin-1 was dissolved in PBS with 0.1% BSA and prepared in 100 μg/ml stock solution. BDNF was dissolved in water and prepared in 50 μg/ml stock solution. Except for (R)-(+)-trans-N-(4-pyridyl)-4-(1-aminoethyl)-cyclohe-xanecarboxamide (Y27632), myristoylated-autocamtid-2-related inhibitory peptide (AIP) and toxin B were dissolved in water, and other chemicals were dissolved in DMSO. All chemicals were prepared in 1000× stock solution and diluted before use.
Results
Ryanodine-induced growth cone turning and local [Ca2+]i elevation
To directly activate an asymmetric elevation of [Ca2+]i across the growth cone in the absence of known guidance factors, we applied an extracellular gradient of ryanodine to the growth cone of cultured Xenopus spinal neurons. Ryanodine is a membrane-permeable ligand that at low concentrations activates Ca2+ release channels in the endoplasmic reticulum (Zucchi and Ronca-Testoni, 1997). Using the standard growth cone turning assay (Lohof et al., 1992; Hong et al., 2000), we observed a marked attractive turning of the growth cone toward the micropipette when the ryanodine was applied (at a pipette concentration of 10 μm) (Fig. 1a). Fluorescence ratio imaging was used to monitor [Ca2+]i under the same turning assay using the ryanodine gradient (see Materials and Methods). Figure 1b shows a line scan of fluorescence ratio (fluo-4/fura-Red) across the growth cone palm coded in pseudocolors before and after the application of the ryanodine gradient. Asymmetric elevation of [Ca2+]i was observed across the growth cone soon after the onset of the ryanodine gradient (at t = 1 min). Transients of [Ca2+]i elevation at the base of filopodia facing the ryanodine pipette were frequently observed (Fig. 1b). To quantitatively assay the [Ca2+]i elevation, we measured changes with time in the average fluorescence ratio on two sides of the growth cone and the growth cone shaft (Fig. 1c, boxes, d). Figure 1e depicts the summary of measurements from 20 growth cones. Significant elevation of [Ca2+]i was observed on the side facing the ryanodine pipette. This ryanodine-induced [Ca2+]i elevation can be prevented by loading a fast Ca2+ buffer BAPTA-AM (10 μm) into these neurons with 30 min preincubation (Fig. 1f). As a control, we loaded the pipette with modified Ringer's solution instead of ryanodine. No elevation of [Ca2+]i was observed across the growth cone using the same assay (Fig. 1f), indicating that the Ca2+ signal observed was not caused by the mechanical perturbation during the turning assay. These observations suggest that a local [Ca2+]i elevation at the proximal side of the growth cone is responsible for ryanodine-induced turning response.

Growth cone turning and local [Ca2+]i elevation induced by ryanodine gradient. a, Microscopic images of a cultured Xenopus spinal neuron at the beginning (0 min) and the end (60 min) at a 1 hexposure to a ryanodine gradient created by pulsatile application of a ryanodine solution (10 μm in the pipette). Superimposed traces (right) depict the trajectory of neurite extension during the 1 h period for all neurons examined. The origin is the center of the growth cone at the onset of the experiment, and the original direction of growth was vertical. The arrow indicates the direction of the gradient. Scale bar, 10 μm. b, Image of a growth cone (top) and line scans of Ca2+ level (bottom) with an interval of 10 ms at the center of the growth cone (red line in the top) before and after the onset of the ryanodine gradient (at t = 1 min). The arrowhead indicates the direction of the ryanodine gradient (10 μm in the pipette). The neuron was loaded with Ca2+-sensitive dyes by incubation with fluo-4 AM (2 μm) and fura-Red AM (2 μm) for 30 min. The line scan shown in b represents the ratio of fluo-4 and fura-Red fluorescence, coded in pseudocolors by a linear scale (shown on the right), with white representing the highest intensity. Traces on the right depict variation of the ratio across the growth cone at different times marked by arrows, normalized to the lowest ratio for each trace. White bar, 10 μm; vertical bar, 50%. c, Image of a growth cone (left) and the pseudocolor-coded fluorescence ratio (fluo-4/fura-Red) before exposure to the ryanodine gradient (right). The arrowhead indicates the direction of the ryanodine gradient. Scale bar, 10 μm. d, The ratio images of the proximal part (1), distal part (2), and shaft (3) of the growth cone at different times are shown at high magnification. e, Summary of normalized changes of fluorescence ratio at different parts of the growth cone before and after exposed to the ryanodine gradient. n refers to the number of growth cones examined. Error bar indicates SEM. f, Similar experiments were performed in e except that modified Ringer's solution was used in the pipette instead of ryanodine-containing solution (top) or that BAPTA-AM (10 μm) was preincubated in bath for 30 min (bottom).
Turning depends on both Ca2+ and Rho GTPases
To further verify that growth cone turning triggered by the ryanodine gradient was indeed caused by [Ca2+]i elevation, rather than other unknown actions of ryanodine, we examined the effect of preventing [Ca2+]i elevation by preincubation of the culture with BAPTA-AM (10 μm) or with thapsigargin (10 μm), which depletes intracellular Ca2+ stores. As shown in Figure 2a, we found that the ryanodine effect was abolished under either condition.
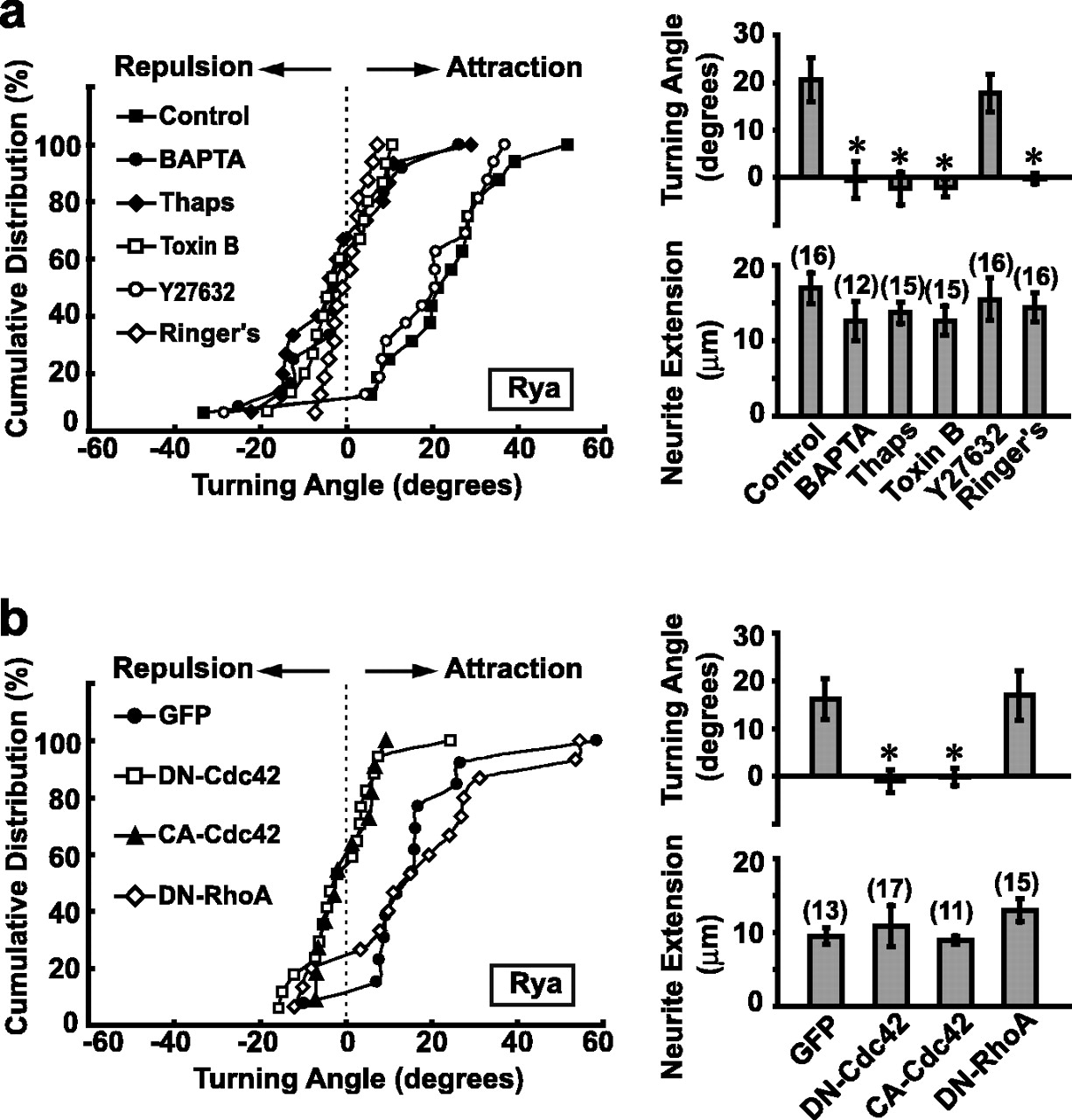
Ryanodine-induced attractive turning response depends on both Ca2+ and Rho GTPases. a, Distribution of growth cone turning angles (left) for control neurons and neurons treated with BAPTA-AM (BAPTA; 10 μm), thapsigargin (Thaps; 10 μm), toxin B (5 ng/ml), or Y27632 (10 μm). Data from the experiment using modified Ringer's solution instead of ryanodine-containing solution were also presented (Ringer's). Histograms (right) represent the average turning angle and neurite extension rate during the 1 h assay for all neurons examined. Asterisk indicates data significantly different from the control (p < 0.01; Kolmogorov-Smirnov test). Error bars indicate SEM. b, Experiments were performed in the same manner as in a, except that the neurons expressed different constructs (GFP alone, DN-Cdc42/GFP, CA-Cdc42/GFP, or DN-RhoA/GFP). Asterisks indicate data significantly different from the GFP control (p < 0.01; Kolmogorov-Smirnov test).
Small GTPases of the Rho family, Cdc42, Rac, and RhoA, have been shown to participate in axon guidance in vivo (Steven et al., 1998; Ng et al., 2002) and to mediate attractive or repulsive growth cone turning induced by a gradient of BDNF or lysophosphatidic acid (LPA), respectively (Yuan et al., 2003). To test whether these Rho GTPases are involved in Ca2+-mediated turning induced by the ryanodine gradient, we first tested toxin B, a specific inhibitor of Rho GTPases, on the ryanodine-induced growth cone turning. As shown in Figure 2a, although many neurites retained substantial growth capacity after overnight treatment of toxin B (at 5 ng/ml), the attractive turning normally induced by the ryanodine gradient was totally abolished. Thus, Rho GTPases are required for ryanodine-induced attractive turning. Furthermore, we found that treatment of the culture with Y27632 (10 μm), a specific inhibitor of Rho kinase, had no effect on ryanodine-induced turning (Fig. 2a), suggesting that that Cdc42/Rac, but not RhoA, is required for the attractive turning.
To examine further the role of Rho family GTPases in mediating the ryanodine-induced attraction, fusion constructs of GFP and mutant Rho GTPases were expressed in Xenopus spinal neurons by the blastomere injection method (Alder et al., 1995). As shown in Figure 2b, a ryanodine gradient (pipette concentration ∼10 μm) caused marked attractive growth cone turning in control neurons or neurons expressing GFP alone. However, the attraction induced by ryanodine was completely abolished in neurons expressing DN or CA form of Cdc42/GFP, but was unaffected in neurons expressing DN-RhoA/GFP (Fig. 2b). These results indicate that Cdc42, but not RhoA, is involved in the attractive turning induced by ryanodine. The effects of Rac mutants and CA-RhoA were not studied in the present work because of severe defects in embryo development and neurite growth caused by the overexpression of these mutants (our unpublished observation), which prevented effective growth cone turning assay (see Materials and Methods).
Ca2+ regulates Rho GTPase activities
The above studies showed that ryanodine-induced growth cone attraction requires both Ca2+ signaling and appropriate Cdc42 activity. Elevation of [Ca2+]i may regulate Rho GTPase activity to trigger growth cone turning. Alternatively, a basal level of Rho GTPase activity may be required as a permissive condition for the Ca2+-dependent turning. We first assayed the effects of extracellular ryanodine on Rho GTPase activity in HEK293T cells transfected with wild-type Rho GTPases, using the affinity binding of active Rho GTPases with their downstream effectors (“pulldown assay”) (Ren et al., 1999) (see Materials and Methods). Cells transfected with wild-type Rho GTPases were treated with different concentrations of ryanodine for 3 min. At a bath concentration of 10 nm ryanodine, which corresponds to approximately that at the growth cone during the attractive turning induced by ryanodine (see Materials and Methods), we observed a transient elevation of both Cdc42 and Rac activities, and a transient reduction of RhoA activity (Fig. 3a,b). The Cdc42 and Rac activities induced by ryanodine (10 nm in bath) reached their peak levels within 3 min and decayed to the basal level by 10 min (Fig. 3b). The modulation of these Rho GTPases was Ca2+ dependent, because preincubation of the cells with BAPTA-AM (10 μm) or thapsigargin (10 μm) for 30 min completely abolished the effect of ryanodine (Fig. 3d). The same concentration of ryanodine also triggered [Ca2+]i elevation in these HEK293T cells (Fig. 3c). This [Ca2+]i elevation was caused by Ca2+ release from internal stores, because no signal was detected when the cells were preincubated for 30 min with thapsigargin (Fig. 3c). In contrast, overnight incubation with toxin B (5 ng/ml) only slightly reduced ryanodine-induced elevation of [Ca2+]i (Fig. 3c).

Ca2+ elevation regulates the activity of Rho GTPases in transfected HEK293T cells. a, b, Representative Western blots showing the dose-response and time course of the activation of Cdc42 and Rac and inactivation of RhoA by bath-applied ryanodine in HEK293T cells transfected with wild-type Rho GTPases. Active GTPases were pulled down by GST-PBD or GST-RBD (see Materials and Methods) and are indicated by asterisks. Protein samples of 1/10th of total lysates were shown to indicate protein loading (bottom). Each data point is the average of three to five independent experiments and represents the activity level normalized by the value of parallel cultures without the ryanodine treatment. Error bars indicate SEM. c, Normalized changes of fluorescence ratio in HEK293T cells were summarized before and after treatment of ryanodine (10 nm) in the presence or absence of thapsigargin (Thaps; 10 μm) or toxin B (5 ng/ml). d, Activation of Cdc42 and inactivation of RhoA by bath-applied ryanodine (10 nm for 3 min) in HEK293T cells transfected with wild-type Rho GTPases were abolished by the pretreatment of BAPTA (10 μm) or thapsigargin (10 μm) for 30 min; n = 3 for each condition.
In primary cultures of cerebellar granule cells, similar Ca2+-dependant regulation of Rho GTPases induced by ryanodine was also observed (Fig. 4a). To further test whether Ca2+ influx through the plasma membrane is similarly effective in regulating Rho GTPases as Ca2+ release from internal stores, we examined the effect of glutamate and a Ca2+ ionophore, A23187, two reagents known to elevate [Ca2+]i. As shown in Figure 4b, at 3 min after bath application of ryanodine (10 nm), glutamate (100 μm) or A23187 (10 μm), we observed a marked upregulation of Cdc42 activity and a downregulation of RhoA activity by the pulldown assay.

Ca2+-dependent regulation of Rho GTPase activity in cultured neurons. a, Activation of Cdc42 and inactivation of RhoA by bath-applied ryanodine (10 nm for 3 min) incultured cerebellar granule cells, assayed by methods as described in Figure 3. The effects of ryanodine on the activity of Rho GTPases were abolished by the pretreatment of BAPTA (10 μm) or thapsigargin (Thaps; 10 μm) for 30 min; n = 3 for each condition. b, The effects of bath-applied ryanodine (10 nm), glutamate (100 μm), and Ca2+ ionophore A23187 (10 μm) on the activity of Cdc42 and RhoA at 3 min after application; n = 4 for each condition. Error bars indicate SEM.
Ca2+-dependent regulation of GTPases by BDNF and netrin-1
We further examined whether there is Ca2+-dependent regulation of Rho GTPases activity by two natural guidance factors, BDNF and netrin-1. Previous studies have shown that a gradient of netrin-1 can trigger a [Ca2+]i gradient in the growth cone (Hong et al., 2000; Henley and Poo, 2004). In the present study, we further studied whether a BDNF gradient can also trigger an asymmetric [Ca2+]i elevation. As shown in Figure 5a, there is a clear elevation of [Ca2+]i at the proximal side of the growth cone facing the BDNF gradient, and the [Ca2+]i elevation is quite lower at the distal side and the shaft of the growth cone. Preincubation of toxin B (5 ng/ml) results in a slight reduction in the rising phase of [Ca2+]i elevation at proximal side of the growth cone (Fig. 5b), consistent with the notion that GTPase activity has a minor effect on [Ca2+]i. We then use the pulldown assay to test whether the Rho GTPase regulation by these natural chemoattractants requires the [Ca2+]i elevation. We found that BDNF (50 ng/ml) can upregulate Cdc42 activity in cultured cerebellar granule cells within 3 min, and this effect was blocked by preincubation with BAPTA-AM (10 μm) or thapsigargin (10 μm) for 30 min (Fig. 5c). Moreover, consistent with a previous report (Li et al., 2002), netrin-1 (100 ng/ml) can upregulate Rac activity in cultured cortical neurons, which express the netrin receptor DCC (deleted in colorectal cancer), and this effect was also blocked by preventing [Ca2+]i elevation with BAPTA-AM (10 μm) or depleting internal Ca2+ store with thapsigargin (10 μm) (Fig. 5d). Thus, activation of Rho GTPases by netrin-1 or BDNF depends on [Ca2+]i elevation, consistent with the model that Rho GTPases mainly function downstream of Ca2+ in axon guidance signaling.

Growth cone [Ca2+]i elevation and Ca2+-dependent regulation of Rho GTPases by BDNF and netrin-1. a, b, Summary of normalized changes of fluorescence ratio at the proximal, distal, and shaft regions of the growth cone before and after exposure to the BDNF gradient in the absence or presence of toxin B (5 ng/ml), presented in the same manner as in Figure 1e. c, Activation of Cdc42 by bath-applied BDNF (50 ng/ml for 3 min) in cultured cerebellar granule cells was abolished by the pretreatment of BAPTA (10 μm) orthapsigargin (Thaps; 10 μm) for 30 min; n = 3. d, Activation of Rac by bath-applied netrin-1 (100 ng/ml for 3 min) in cultured cortical neurons was abolished by the pretreatment of BAPTA (10 μm) or thapsigargin (10 μm) for 30 min; n = 4. Error bars indicate SEM.
PKC mediates Ca2+ signaling
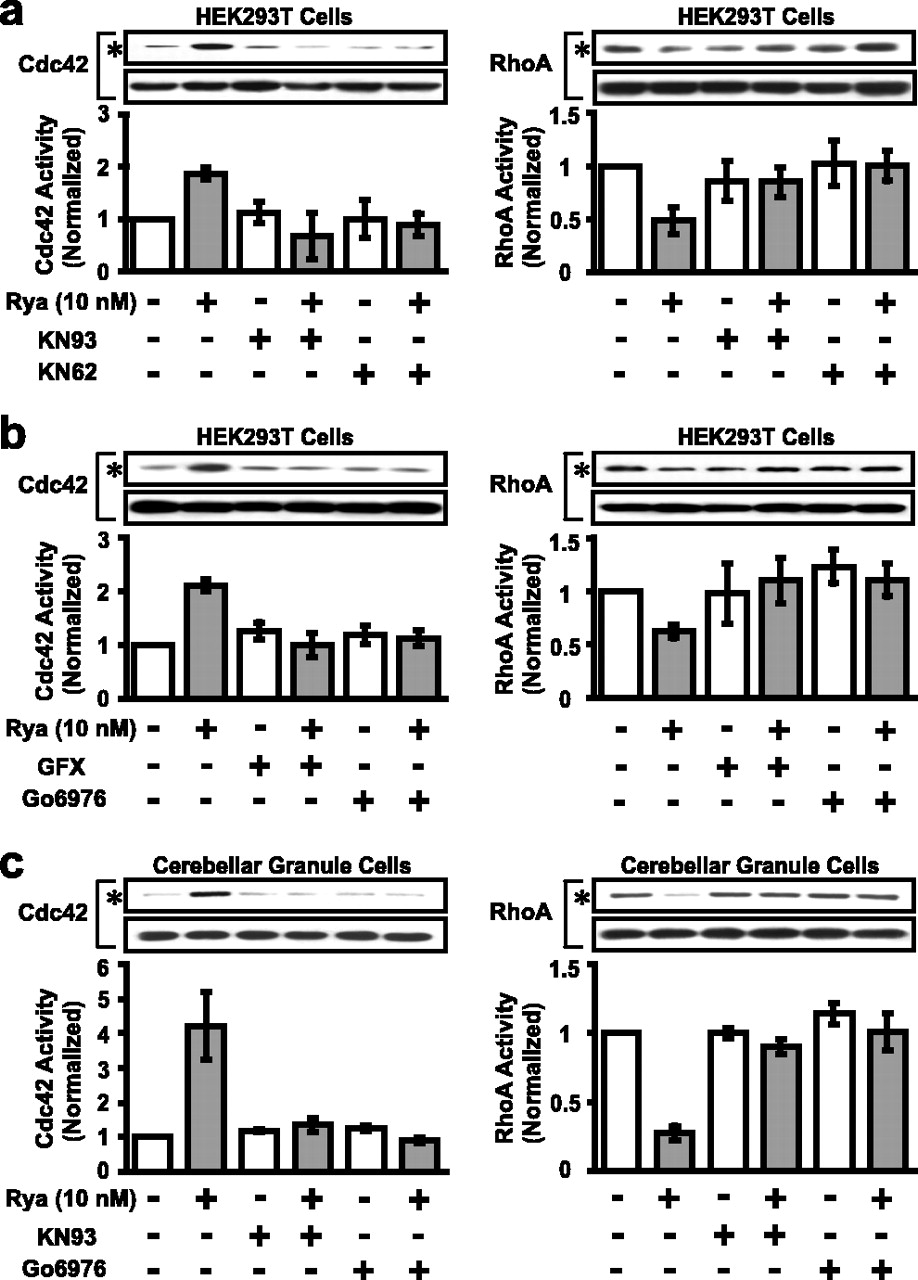
The above studies suggest that Ca2+ signaling lies at upstream of Rho GTPases. We next examined potential effectors of Ca2+ that may act on these GTPases. As shown in Figure 6, a and b, we found that preincubation with drugs that inhibit CaMKII (KN93, 2.5 μm; KN62, 2.5 μm) or PKC [2-[1-(3-dimethylaminopropyl)-1H-indol-3-yl]-3-(1H-indol-3-yl)-maleimide (GFX), 50 nm; 12-(2-cyanoethyl)-6,7,12,13-tetrahydro-13-methyl-5-oxo-5H-in-dolo(2,3-a) pyrrolo(3,4-c)-carbazole (Go6976), 50 nm] respectively, all dramatically reduced ryanodine-induced activation of Cdc42 and inhibition of RhoA in HEK293T cells expressing these proteins. In cultured cerebellar granule cells, similar effects were observed that Rho GTPase regulation by ryanodine can be blocked by inhibition of either PKC or CaMKII (Fig. 6c). These results are consistent with previous reports indicating that CaMKII and PKC can regulate the activity of Rho GTPases (Fleming et al., 1999; Price et al., 2003). Treatment of Xenopus spinal neurons with these inhibitors also blocked the growth cone turning of induced by the ryanodine or netrin-1 gradient (Fig. 7a,b). The blockade of netrin-1-induced growth cone attraction is consistent with a recent report that a CaMKII/calcineurin switch controls the direction of Ca2+-dependent growth cone guidance (Wen et al., 2004). In contrast, inhibition of Rho kinase (by Y27632, 10 μm), PI-3 kinase [by 2-(4-morpholinyl)-8-phenyl-1-(4H)-benzopyran-4-one (LY294002), 10 μm] protein phosphatase I (by caliculin A, 0.5 nm), or myosin light chain kinase [by 1-(5-iodonaphthalene-1-sulfonyl)-1H-hexahydro-1,4-diazepine HCl (ML-7), 5 μm]did not affect growth cone attraction triggered by the ryanodine gradient (Fig. 7c).

Regulation of the activities of Rho GTPases by ryanodine requires CaMKII and PKC. a, b, Regulation of Rho GTPases by bath-applied ryanodine (10 nm for 3 min) in HEK293T cells transfected with wild-type Rho GTPases was abolished by specific inhibitors of CaMKII (KN93 or KN62, 2.5 μm each; n = 6 for Cdc42; n = 4 for RhoA) or PKC (GFX or Go6976, 50 nm each; n = 8 for Cdc42; n = 5 for RhoA). c, Regulation of Rho GTPases by bath-applied ryanodine (10 nm for 3 min) in cultured cerebellar granule cells was also abolished by KN93 (2.5 μm) or Go6976 (50 nm); n = 4 for Cdc42 and n = 3 for RhoA. Error bars indicate SEM.

Ryanodine-induced attraction depends on CaMKII and PKC. Distribution of turning angles in a gradient of ryanodine (a, c, 10 μm in the pipette) or netrin-1 (b, 5 μg/ml in the pipette) for cultured Xenopus spinal neurons in control condition or after treatment with various inhibitors: KN62 (2.5 μm), AIP (200 nm), GFX (50 nm), ML-7 (5 μm), LY294002 (LY; 10 μm), and caliculin A (CLCA; 0.5 nm). Histograms represent the average turning angle and neurite extension rate during the 1 h assay for these experiments. Asterisk indicates data significantly different from the control (p < 0.01; Kolmogorov-Smirnov test). Error bars indicate SEM.
The above results suggest that ryanodine-induced Ca2+ signals may activate PKC and CaMKII to regulate Rho GTPases. Alternatively, a basal activity of these enzymes in the cell is necessary for regulating Rho GTPases. To distinguish these two possibilities, we further measured the activity of these enzymes in response to the ryanodine treatment in both HEK293T cells and neurons. Western blotting was used to detect the active PKCα/β and CaMKII in the cell lysates, using phosphospecific antibodies specific to PKCα/β and CaMKII, respectively. As shown in Figure 8, a and b, we found a rapid increase of PKCα/β activity both in HEK293T cells and in cultured cerebellar granule cells in response to ryanodine treatment (10 nm), but no obvious change in the CaMKII activity. Moreover, in cultures of embryonic cortical neurons, which express the netrin receptor DCC, PKC was also transiently activated by netrin-1 without changing CaMKII activity (our unpublished observation). Together, these results suggest that the Ca2+ signaling to Rho GTPases and growth cone turning is mediated by PKCα/β, whereas a basal activity of CaMKII is required as a permissive condition for Ca2+-dependent Rho GTPase regulation.

PKC mediates Ca2+ signaling to Rho GTPases. a, b, Western blots showing the active form of PKC and CaMKII in HEK293T cells or cultured cerebellar granule cells treated with ryanodine (10 nm) at different times. Western blots using antibody against actin (bottom) are shown as the loading control. Graphs below depict average kinase activity normalized to values obtained from cells not treated with ryanodine; n = 4 for each condition. Error bars indicate SEM. c, Microscopic images of a cultured Xenopus spinal neuron at the beginning and the end of a 1 h exposure to a gradient of KN62 or Go6976 (2.5 mm or 50 μm in the pipette). Superimposed traces depict the trajectory of neurite extension during the 1 h period for all neurons examined. Scale bar, 10 μm. d, Distribution of turning angles (left) for neurons in a gradient of KN62 or Go6976. Histograms (right) represent the average turning angle and neurite extension under various conditions including overnight treatment of toxin B and using Ringer's solution in the pipette. Asterisk indicates data significantly different from Ringer's control (p < 0.01; Kolmogorov-Smirnov test). Error bars indicate SEM.
To further test the role of these enzymes in growth cone turning, we applied an extracellular gradient of enzyme inhibitors, which can presumably generate a gradient of enzyme activity across the growth cone, with lower enzyme activity on the side facing the gradient. We found that a gradient of Go6976 (at both 50 and 10 μm in the pipette), induced repulsive turning (Fig. 8c,d), which is consistent with the previous report that a micropipette containing the PKC activator 12-O-tetradecanoyl-phorbol 13-acetate or phorbol 12,13-dibutyrate, produced a significant growth cone attraction (Fu et al., 1997). In contrast, a gradient of KN62 (2.5 and 0.5 mm in the pipette) or KN93 (2.5 and 0.5 mm in the pipette) did not induce significant turning responses (Fig. 8c,d). The repulsion induced by the activity gradient of PKCα/β also requires Rho GTPase activity, because overnight preincubation with toxin B (5 ng/ml) totally abolished the repulsive effect of the Go6976 gradient (Fig. 8c,d). Thus, an activity gradient of PKCα/β across the growth cone can indeed mediate growth cone turning signal through Rho GTPases, driving growth cone toward the side with higher enzyme activity.
Discussion
A gradient of [Ca2+]i was reported to mediate the growth cone turning signaling (Hong et al., 2000; Zheng, 2000). In the present study, we observed a distinct asymmetric [Ca2+]i elevation in the growth cone after exposure to an extracellular gradient of ryanodine (Fig. 1b-e). Although previous analysis predicted a concentration difference across the growth cone to be only 10% of the average concentration at the growth cone (Lohof et al., 1992; Zheng et al., 1994), the highly localized [Ca2+]i elevation on the side facing the ryanodine gradient may be related to nonlinearity associated with Ca2+-induced Ca2+ release. Gradient amplification is known to occur and is required for cellular detection of small extracellular gradient signals, as exemplified by many polarized cell motility (Servant et al., 2000; Levchenko and Iglesias, 2002). An autocatalytic amplification process at the growth cone that enhances local Ca2+ release/accumulation and inhibits Ca2+ release at a more distant site may serve to amplify the localized signal. The detailed mechanism for the amplification of [Ca2+]i gradient at the growth cone remains to be clarified.
Both Ca2+ and Rho GTPases are known to play essential roles in axon guidance in various systems. In cultured Xenopus spinal neurons, the turning response induced by a gradient of BDNF was blocked by either perturbing Cdc42 activity (Yuan et al., 2003) or preventing [Ca2+]i elevation (Song and Poo, 1999, 2001). In the present study, we investigated the causal relationship between these two signaling events. We found that the local [Ca2+]i elevation in the growth cone drives the axon toward the higher [Ca2+]i side. Because perturbing the gradient of Cdc42 activity by uniformly expressing either DN-Cdc42 or CA-Cdc42 blocked this turning (Fig. 2), and [Ca2+]i elevation can activate Cdc42 and Rac (Figs. 3, 4), Cdc42 appears to be a downstream effector of Ca2+ that mediates the growth cone turning. Although [Ca2+]i elevation can also decrease RhoA activity (Fig. 3), RhoA does not play essential roles in this [Ca2+]i-triggered attraction, because neither DN-RhoA nor Y27632 [the specific ROCK (Rho-associated protein kinase) inhibitor] affected ryanodine-triggered growth cone attraction (Fig. 2). The effect of mutated Rac proteins on the turning response was not tested in this study because of technical difficulties. Based on our biochemical assay of Rac activity, we suggest that Rac can be activated by [Ca2+]i and acts in parallel with Cdc42 in mediating growth cone turning, consistent with the previous finding that these two GTPases share many common upstream regulators and downstream effectors (Mackay and Hall, 1998).
In the present study, we found in cultured cerebellar granule cells that Ca2+ influx triggered by A23187 or glutamate is similarly effective in regulating Rho GTPases as ryanodine-induced Ca2+ release from internal stores (Fig. 4b,c), suggesting that the effect may be independent of the Ca2+ sources in our culture preparation. Consistently, attractive turning of Xenopus growth cones can be triggered by a gradient of either ryanodine (Fig. 1a), glutamate (Zheng et al., 1996), or A23187 (Henley and Poo, 2004).
It has been shown that Rho GTPases can also regulate the Ca2+ dynamics in the cytosol (Wilk-Blaszczak et al., 1997; Costello et al., 1999; Djouder et al., 2000; Singleton and Bourguignon, 2002). We found that toxin B treatment caused a slight reduction in the rising phase of the [Ca2+]i elevation in response to BDNF (Fig. 5b), consistent with the notion of a feedback regulation from Rho GTPases to Ca2+ signaling during axon guidance. However, based on the following observations we propose that Cdc42/Rac acts mainly downstream of Ca2+ signal during growth cone turning. First, after toxin B treatment, significant asymmetric [Ca2+]i elevation can be triggered by BDNF in Xenopus growth cones (Fig. 5b); Second, the growth cone turning triggered by the activity gradient of Ca2+ effector enzyme PKC also requires Rho GTPases activities (Fig. 8d). Together, our data support the notion that Cdc42/Rac mainly acts downstream of Ca2+ signals in mediating growth cone turning and a gradient of [Ca2+]i in the growth cone leads to a gradient of Cdc42/Rac activity, which in turn provides the directional cue for cytoskeletal rearrangement (supplemental Fig. 1, available at www.jneurosci.org as supplemental material). In addition, Rho GTPases may also speed up the formation of [Ca2+]i gradient as a positive feedback to Ca2+ signals in triggering growth cone turning.
Previous studies have shown that Rho GTPases can mediate axon guidance by BDNF (Yuan et al., 2003) and that BDNF effector enzymes phospholipase C γ (PLCγ) and phosphatidylinositol 3-kinase may regulate Rho GTPase activity through Ca2+-dependent and Ca2+-independent mechanisms, respectively (Fleming et al., 1998; Akasaki et al., 1999). The present study shows that the BDNF-induced Cdc42 activity and the netrin-induced Rac activity are Ca2+ dependent, because they can be abolished by BAPTA-AM or thapsigargin (Fig. 5c,d). Together with previous results, we suggest that a BDNF or netrin-1 gradient triggers the [Ca2+]i gradient in the growth cone (supplemental Fig. 1, available at www.jneurosci.org as supplemental material), which in turn creates a gradient of Rho GTPase activity to drive the turning response.
Other guidance molecules may also activate Rho GTPases through signaling processes that have not been shown to be linked to Ca2+. For example, slit was shown to increase srGAP1-Robo1 interaction and inactivate Cdc42 in subventricular zone neurons (Wong et al., 2001). Semaphorin also activates plexin, leading to the upregulation of postsynaptic density-95/discs large/zona occludens-1-RhoGEF/leukemia-associated RhoGEF and RhoA activity in hippocampal neurons (Aurandt et al., 2002; Perrot et al., 2002; Swiercz et al., 2002). However, whether Ca2+ is a coregulator during these processes remains to be examined. Because all these axon guidance factors trigger both Ca2+ and Rho GTPase signaling, three possibilities exist: (1) Ca2+ acts upstream of Rho GTPases, as shown in the present study, (2) [Ca2+]i elevation is required as a coactivator of some GTP exchange factors (GEFs) or GTPase-activating proteins (GAPs) for the Rho GTPase regulation, (3) Ca2+ and Rho GTPases are independently activated and are both required for axon guidance. It is reasonable to speculate that these three modes of Ca2+ actions contribute to different extent in axon guidance signaling by different factors.
Ca2+-dependent regulation of Rho GTPases has been well documented in non-neuronal cells (Fleming et al., 1998, 1999; Price et al., 2003), although how changes in [Ca2+]i affect these GTPases activity is essentially unclear. In PC3 human prostate carcinoma cells, increased [Ca2+]i activates conventional PKC, which phosphorylates RhoGDIα and induces the translocation and activation of Rac (Price et al., 2003). There is also evidence in Swiss 3T3 fibroblasts that PLCγ, PKC, and CaMKII are involved in the phosphorylation of the Rac GEF Tiam1 induced by platelet-derived growth factor (Fleming et al., 1998, 1999). Thus, Ca2+-dependent regulation of Rho GTPases may also be mediated by phosphorylation of GEFs, GAPs, or GDP dissociation inhibitors in neurons. In the present study, we found that inhibition of either CaMKII or PKC can abolish the regulation of Rho GTPases activity by ryanodine in both HEK293T cells and neurons (Fig. 6). Because PKC, but not CaMKII, is activated by ryanodine (10 nm) in the neuronal cells (Fig. 8b), we propose that PKC is responsible for the ryanodine-induced upregulation of Cdc42/Rac, possibly through phosphorylation of Tiam1 or other regulators, whereas a basal level of CaMKII activity is required for but does not mediate the regulation of Rho GTPases by ryanodine (supplemental Fig. 1, available at www.jneurosci.org as supplemental material). There is evidence that GEFs and GAPs participate in axon guidance in vivo (Dickson, 2002). It remains to be elucidated which ones of these regulatory proteins of GTPases can be activated or inhibited by Ca2+-dependent PKC in response to guidance factors.
PKC can mediate neurite guidance at the astrocyte boundary in coculture of neurons and astrocytes (Powell et al., 2001) and participate in the midline axon guidance in mice through its phosphorylation of GAP-43 (Zhang et al., 2000; Shen et al., 2002). A gradient of PKC activator has been reported to produce a significant growth cone attraction (Fu et al., 1997). Here we found that Ca2+-dependent PKC, which can be rapidly activated by ryanodine (Fig. 8a,b), is required for netrin- and ryanodine-triggered growth cone attraction (Fig. 7) and is responsible for the Rho GTPase regulation by ryanodine (Fig. 6). Thus Ca2+-dependent PKC may be rapidly and locally regulated and contribute to chemoattraction of the growth cone through its local upregulation of Cdc42/Rac, which, in turn, promotes growth cone filopodia elongation as well as microtubule stabilization (Cau et al., 2001; Daub et al., 2001).
In summary, we have shown in the present study that an asymmetric [Ca2+]i elevation across the growth cone is sufficient to drive the growth cone turning toward the higher [Ca2+]i side, and such turning requires appropriate Cdc42 activity. Direct elevation of [Ca2+]i in neurons can transiently activate Cdc42/Rac, but inactivate RhoA. Regulation of Rho GTPases by BDNF and netrin-1 also requires [Ca2+]i elevation. The signaling from Ca2+ to Cdc42 is mediated by PKC and requires a basal activity of CaMKII. These findings provide direct evidence that Rho GTPases can act downstream of Ca2+ signals to trigger growth cone turning. They also support the notion that a gradient of Rho GTPase activity induced by [Ca2+]i gradient across the growth cone may directly mediate axon guidance.
Footnotes
This work was supported by grants from the Major State Basic Research Program of China and the Shanghai Science and Technology Development Foundation. X.-b.Y. was supported by the National Science Foundation (30300103) of China and by the Rising Star Project of Shanghai (2003). M.-m.P. was supported in part by National Institutes of Health Grant NS 22764. We thank L. Mei, D. Turner, A. Hall, and G. Bokoch for providing cDNA clones and X. Xu for cDNA blastomere injection.
Correspondence should be addressed to either of the following: M. Poo, Division of Neurobiology, Department of Molecular and Cell Biology, University of California, Berkeley, CA 94720-3200, E-mail: mpoo{at}uclink.berkeley.edu; or X. Yuan, Institute of Neuroscience, Shanghai Institutes of Biological Sciences, Chinese Academy of Sciences, Shanghai 200031, China, E-mail: yuanxb{at}ion.ac.cn.
Y.-q. Song's present address: Institute of Neuroscience, University of Pennsylvania, Philadelphia, PA 19104.
Copyright © 2005 Society for Neuroscience 0270-6474/05/252338-10$15.00/0
↵* M.J., C.-b.G., and Y.-a.J. contributed equally to this work.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}