
Abstract
Immunotherapies that produce durable responses in some malignancies have failed in pancreatic ductal adenocarcinoma (PDAC) due to rampant immune suppression and poor tumor immunogenicity. We and others have demonstrated that induction of the senescence-associated secretory phenotype (SASP) can be an effective approach to activate anti-tumor natural killer (NK) cell and T cell immunity. In the present study, we found that the pancreas tumor microenvironment suppresses NK cell and T cell surveillance after therapy-induced senescence through enhancer of zeste homolog 2 (EZH2)-mediated epigenetic repression of proinflammatory SASP genes. EZH2 blockade stimulated production of SASP chemokines CCL2 and CXCL9/10, leading to enhanced NK cell and T cell infiltration and PDAC eradication in mouse models. EZH2 activity was also associated with suppression of chemokine signaling and cytotoxic lymphocytes and reduced survival in patients with PDAC. These results demonstrate that EZH2 represses the proinflammatory SASP and that EZH2 inhibition combined with senescence-inducing therapy could be a powerful means to achieve immune-mediated tumor control in PDAC.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others
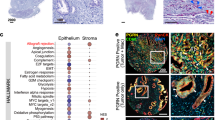
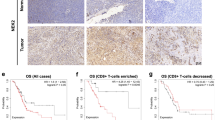
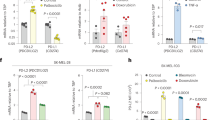
Data availability
RNA-seq and CUT&Tag data that support the findings of the present study have been deposited in the GEO under accession nos. GSE141684, GSE201495 and GSE203623. Datasets derived from this resource that support the findings of the present study are available in Supplementary Tables 1–46. Gene expression data for human LUAD and PDAC cell lines treated with T/P were obtained under accession no. GSE110397. Gene expression data from 145 primary human PDAC specimens were obtained under accession no. GSE71729. Source data are provided with this paper. All other data supporting the findings of the present study are available from the corresponding author upon reasonable request.
References
Siegel, R. L., Miller, K. D., Fuchs, H. E. & Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 72, 7–33 (2022).
Olive, K. P. et al. Inhibition of Hedgehog signaling enhances delivery of chemotherapy in a mouse model of pancreatic cancer. Science 324, 1457–1461 (2009).
Provenzano, P. P. et al. Enzymatic targeting of the stroma ablates physical barriers to treatment of pancreatic ductal adenocarcinoma. Cancer Cell 21, 418–429 (2012).
Brahmer, J. R. et al. Safety and activity of anti-PD-L1 antibody in patients with advanced cancer. N. Engl. J. Med. 366, 2455–2465 (2012).
Royal, R. E. et al. Phase 2 trial of single agent Ipilimumab (anti-CTLA-4) for locally advanced or metastatic pancreatic adenocarcinoma. J. Immunother. 33, 828–833 (2010).
Haas, A. R. et al. Phase I study of lentiviral-transduced chimeric antigen receptor-modified T cells recognizing mesothelin in advanced solid cancers. Mol. Ther. 27, 1919–1929 (2019).
Bear, A. S., Vonderheide, R. H. & O’Hara, M. H. Challenges and opportunities for pancreatic cancer immunotherapy. Cancer Cell 38, 788–802 (2020).
Waters, A. M. & Der, C. J. KRAS: the critical driver and therapeutic target for pancreatic cancer. Cold Spring Harb. Perspect. Med. https://doi.org/10.1101/cshperspect.a031435 (2018).
Nguyen, K. B. & Spranger, S. Modulation of the immune microenvironment by tumor-intrinsic oncogenic signaling. J. Cell Biol. https://doi.org/10.1083/jcb.201908224 (2020).
Ischenko, I. et al. KRAS drives immune evasion in a genetic model of pancreatic cancer. Nat. Commun. 12, 1482 (2021).
Hamarsheh, S., Gross, O., Brummer, T. & Zeiser, R. Immune modulatory effects of oncogenic KRAS in cancer. Nat. Commun. 11, 5439 (2020).
Canon, J. et al. The clinical KRAS(G12C) inhibitor AMG 510 drives anti-tumour immunity. Nature 575, 217–223 (2019).
Ribas, A. et al. Combined BRAF and MEK inhibition with PD-1 blockade immunotherapy in BRAF-mutant melanoma. Nat. Med. 25, 936–940 (2019).
Ebert, P. J. R. et al. MAP kinase inhibition promotes T cell and anti-tumor activity in combination with PD-L1 checkpoint blockade. Immunity 44, 609–621 (2016).
Ruscetti, M. et al. NK cell-mediated cytotoxicity contributes to tumor control by a cytostatic drug combination. Science 362, 1416–1422 (2018).
Ruscetti, M. et al. Senescence-induced vascular remodeling creates therapeutic vulnerabilities in pancreas cancer. Cell 181, 424–441.e421 (2020).
Faget, D. V., Ren, Q. & Stewart, S. A. Unmasking senescence: context-dependent effects of SASP in cancer. Nat. Rev. Cancer 19, 439–453 (2019).
Coppe, J. P. et al. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 6, 2853–2868 (2008).
Vilgelm, A. E. et al. Connecting the dots: therapy-induced senescence and a tumor-suppressive immune microenvironment. J. Natl Cancer Inst. 108, djv406 (2016).
Kang, T. W. et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature 479, 547–551 (2011).
Xue, W. et al. Senescence and tumour clearance is triggered by p53 restoration in murine liver carcinomas. Nature 445, 656–660 (2007).
Chibaya, L., Snyder, J. & Ruscetti, M. Senescence and the tumor-immune landscape: implications for cancer immunotherapy. Semin. Cancer Biol. https://doi.org/10.1016/j.semcancer.2022.02.005 (2022).
Antonangeli, F., Zingoni, A., Soriani, A. & Santoni, A. Senescent cells: living or dying is a matter of NK cells. J. Leukoc. Biol. 105, 1275–1283 (2019).
Huntington, N. D., Cursons, J. & Rautela, J. The cancer-natural killer cell immunity cycle. Nat. Rev. Cancer 20, 437–454 (2020).
Hingorani, S. R. et al. Preinvasive and invasive ductal pancreatic cancer and its early detection in the mouse. Cancer Cell 4, 437–450 (2003).
Jackson, E. L. et al. Analysis of lung tumor initiation and progression using conditional expression of oncogenic K-ras. Genes Dev. 15, 3243–3248 (2001).
Serrano, M., Lin, A. W., McCurrach, M. E., Beach, D. & Lowe, S. W. Oncogenic ras provokes premature cell senescence associated with accumulation of p53 and p16INK4a. Cell 88, 593–602 (1997).
Narita, M. et al. Rb-mediated heterochromatin formation and silencing of E2F target genes during cellular senescence. Cell 113, 703–716 (2003).
Chien, Y. et al. Control of the senescence-associated secretory phenotype by NF-kappaB promotes senescence and enhances chemosensitivity. Genes Dev. 25, 2125–2136 (2011).
Gluck, S. et al. Innate immune sensing of cytosolic chromatin fragments through cGAS promotes senescence. Nat. Cell Biol. 19, 1061–1070 (2017).
Hoare, M. et al. NOTCH1 mediates a switch between two distinct secretomes during senescence. Nat. Cell Biol. 18, 979–992 (2016).
Tasdemir, N. et al. BRD4 connects enhancer remodeling to senescence immune surveillance. Cancer Discov. 6, 612–629 (2016).
Laugesen, A., Hojfeldt, J. W. & Helin, K. Role of the polycomb repressive complex 2 (PRC2) in transcriptional regulation and cancer. Cold Spring Harb. Perspect. Med. https://doi.org/10.1101/cshperspect.a026575 (2016).
Ito, T., Teo, Y. V., Evans, S. A., Neretti, N. & Sedivy, J. M. Regulation of cellular senescence by polycomb chromatin modifiers through distinct DNA damage- and histone methylation-dependent pathways. Cell Rep. 22, 3480–3492 (2018).
Bracken, A. P. et al. The polycomb group proteins bind throughout the INK4A-ARF locus and are disassociated in senescent cells. Genes Dev. 21, 525–530 (2007).
Yasuda, T. et al. Inflammation-driven senescence-associated secretory phenotype in cancer-associated fibroblasts enhances peritoneal dissemination. Cell Rep. 34, 108779 (2021).
Whittle, M. C. & Hingorani, S. R. Fibroblasts in pancreatic ductal adenocarcinoma: biological mechanisms and therapeutic targets. Gastroenterology 156, 2085–2096 (2019).
Ozdemir, B. C. et al. Depletion of carcinoma-associated fibroblasts and fibrosis induces immunosuppression and accelerates pancreas cancer with reduced survival. Cancer Cell 25, 719–734 (2014).
Gan, L. et al. Epigenetic regulation of cancer progression by EZH2: from biological insights to therapeutic potential. Biomark. Res. 6, 10 (2018).
Iannello, A., Thompson, T. W., Ardolino, M., Lowe, S. W. & Raulet, D. H. p53-dependent chemokine production by senescent tumor cells supports NKG2D-dependent tumor elimination by natural killer cells. J. Exp. Med. 210, 2057–2069 (2013).
Litchfield, K. et al. Meta-analysis of tumor- and T cell-intrinsic mechanisms of sensitization to checkpoint inhibition. Cell 184, 596–614.e514 (2021).
Chow, M. T. et al. Intratumoral activity of the CXCR3 chemokine system is required for the efficacy of anti-PD-1 therapy. Immunity 50, 1498–1512.e1495 (2019).
Reschke, R. et al. Immune cell and tumor cell-derived CXCL10 is indicative of immunotherapy response in metastatic melanoma. J. Immunother. Cancer https://doi.org/10.1136/jitc-2021-003521 (2021).
Moffitt, R. A. et al. Virtual microdissection identifies distinct tumor- and stroma-specific subtypes of pancreatic ductal adenocarcinoma. Nat. Genet. 47, 1168–1178 (2015).
Ho, W. J., Jaffee, E. M. & Zheng, L. The tumour microenvironment in pancreatic cancer—clinical challenges and opportunities. Nat. Rev. Clin. Oncol. 17, 527–540 (2020).
Peng, D. et al. Epigenetic silencing of TH1-type chemokines shapes tumour immunity and immunotherapy. Nature 527, 249–253 (2015).
Zingg, D. et al. The histone methyltransferase Ezh2 controls mechanisms of adaptive resistance to tumor immunotherapy. Cell Rep. 20, 854–867 (2017).
Bugide, S., Green, M. R. & Wajapeyee, N. Inhibition of enhancer of zeste homolog 2 (EZH2) induces natural killer cell-mediated eradication of hepatocellular carcinoma cells. Proc. Natl Acad. Sci. USA 115, E3509–E3518 (2018).
Bugide, S., Gupta, R., Green, M. R. & Wajapeyee, N. EZH2 inhibits NK cell-mediated antitumor immunity by suppressing CXCL10 expression in an HDAC10-dependent manner. Proc. Natl Acad. Sci. USA https://doi.org/10.1073/pnas.2102718118 (2021).
Wang, J. & Wang, G. G. No easy way out for EZH2: its pleiotropic, noncanonical effects on gene regulation and cellular function. Int. J. Mol. Sci. https://doi.org/10.3390/ijms21249501 (2020).
Ougolkov, A. V., Bilim, V. N. & Billadeau, D. D. Regulation of pancreatic tumor cell proliferation and chemoresistance by the histone methyltransferase enhancer of zeste homologue 2. Clin. Cancer Res. 14, 6790–6796 (2008).
Patil, S. et al. EZH2 regulates pancreatic cancer subtype identity and tumor progression via transcriptional repression of GATA6. Cancer Res. 80, 4620–4632 (2020).
Ghosh, M. et al. Mutant p53 suppresses innate immune signaling to promote tumorigenesis. Cancer Cell 39, 494–508 e495 (2021).
Shao, F. F., Chen, B. J. & Wu, G. Q. The functions of EZH2 in immune cells: principles for novel immunotherapies. J. Leukoc. Biol. 110, 77–87 (2021).
Kim, W. et al. Targeted disruption of the EZH2-EED complex inhibits EZH2-dependent cancer. Nat. Chem. Biol. 9, 643–650 (2013).
Tomassi, S., Romanelli, A., Zwergel, C., Valente, S. & Mai, A. Polycomb repressive complex 2 modulation through the development of EZH2–EED interaction inhibitors and EED binders. J. Med. Chem. 64, 11774–11797 (2021).
Ma, A. et al. Discovery of a first-in-class EZH2 selective degrader. Nat. Chem. Biol. 16, 214–222 (2020).
Wang, J. et al. EZH2 noncanonically binds cMyc and p300 through a cryptic transactivation domain to mediate gene activation and promote oncogenesis. Nat. Cell Biol. 24, 384–399 (2022).
Li, C. et al. Finding an easy way to harmonize: a review of advances in clinical research and combination strategies of EZH2 inhibitors. Clin. Epigenet. 13, 62 (2021).
Ram, S. et al. Pixelwise H-score: a novel digital image analysis-based metric to quantify membrane biomarker expression from immunohistochemistry images. PLoS ONE 16, e0245638 (2021).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Yu, G., Wang, L.-G., Han, Y. & He, Q.-Y. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS 16, 284–287 (2012).
Leek, J. T. svaseq: removing batch effects and other unwanted noise from sequencing data. Nucleic Acids Res. https://doi.org/10.1093/nar/gku864 (2014).
Chen, E. Y. et al. Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinf. 14, 128 (2013).
Kaya-Okur, H. S. et al. CUT&Tag for efficient epigenomic profiling of small samples and single cells. Nat. Commun. 10, 1930 (2019).
Gopalan, S., Wang, Y., Harper, N. W., Garber, M. & Fazzio, T. G. Simultaneous profiling of multiple chromatin proteins in the same cells. Mol. Cell 81, 4736–4746.e4735 (2021).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Li, H. et al. The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Ramírez, F., Dündar, F., Diehl, S., Grüning, B. A. & Manke, T. deepTools: a flexible platform for exploring deep-sequencing data. Nucleic Acids Res. 42, W187–W191 (2014).
Zerbino, D. R., Johnson, N., Juettemann, T., Wilder, S. P. & Flicek, P. WiggleTools: parallel processing of large collections of genome-wide datasets for visualization and statistical analysis. Bioinformatics 30, 1008–1009 (2014).
Ross-Innes, C. S. et al. Differential oestrogen receptor binding is associated with clinical outcome in breast cancer. Nature 481, 389–393 (2012).
Yu, J. et al. A polycomb repression signature in metastatic prostate cancer predicts cancer outcome. Cancer Res. 67, 10657–10663 (2007).
Cursons, J. et al. A gene signature predicting natural killer cell infiltration and improved survival in melanoma patients. Cancer Immunol. Res. 7, 1162–1174 (2019).
Cozar, B. et al. Tumor-infiltrating natural killer cells. Cancer Discov. 11, 34–44 (2021).
Acknowledgements
We thank K. Hatzi for providing shRNA constructs, R. Mezzadra for generating Ccl2 O/E cell lines, G. Cottle for technical assistance, C. Baer and C. Hung in the Sanderson Center for Optical Experimentation (SCOPE) facility (RRID:SCR_022721) and Y. Liu in the Morphology Core at UMass Chan Medical School for assistance with IHC analysis and quantification, J. Pitarresi for assistance with coimmunofluorescence staining and analysis, and W. Xue, A. Mercurio, M. Kelliher, M. Green, J. Chuprin, L. Zhou and other members of the Ruscetti laboratory for helpful suggestions and comments on the manuscript. Graphics in Figs. 1a–c, 2a, 3a and 5a were created with BioRender.com. This work was supported by a K99/R00 CA241110 grant from the National Cancer Institute to M.R. and a Memorial Sloan Kettering Cancer Center Support grant (no. P30 CA008748) to S.W.L. We acknowledge support from Our Danny Cancer Fund (no. U6035343000000W to L.C.), the National Institutes of Health (grant nos. R01 HD072122 to T.G.F. and P30 CA008748 S5 to E.d.S.) and the National Center for Advancing Translational Sciences (grant no. UL1-TR001453 to K.S.). S.W.L. is the Geoffrey Beene Chair for Cancer Biology and a Howard Hughes Medical Institute investigator.
Author information
Authors and Affiliations
Contributions
L.C. and M.R. conceived the study, designed and performed experiments, interpreted results and wrote the paper with assistance from all authors. K.C.M., K.D.D., Y.L.-D., J.P.M. and S.C. designed, performed and analyzed in vitro and in vivo experiments. K.C.M., K.D.D., C.N.P., Y.L.-D., J.S., W.L., A.K. and E.d.S. produced and treated animal models. H.L., J.L., Y.-j.H. and L.J.Z. analyzed transcriptomic datasets. S.G., T.G.F., H.L. and L.J.Z. designed and performed CUT&Tag analysis. M.F. and K.S. provided human PDAC patient specimens and data. M.R. and S.W.L. supervised the study.
Corresponding authors
Ethics declarations
Competing interests
S.W.L. is a founder and member of the scientific advisory board of Blueprint Medicines, Mirimus Inc., ORIC Pharmaceuticals, Senescea and Faeth Therapeutics, is on the scientific advisory board of PMV Pharmaceuticals, and is a consultant for Fate Therapeutics. M.R. is a consultant for Boehringer Ingelheim. L.C. and M.R. have filed a US patent application (US22/45163) related to this work. The remaining authors declare no competing interests.
Peer review
Peer review information
Nature Cancer thanks Manuel Collado and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 T/P treatment induces cellular senescence across tumor conditions in vivo.
a, Representative Hematoxylin and eosin (H&E) (top) and Masson’s trichrome (bottom) staining of indicated KPC1 PDAC (PIP, PIL, PILiver) and KP1 LUAD (LIL, LIP, LILiver) derived-tumors grown in different organs from 2-3 independent experiments. Scale bars, 100μm. b, Immunohistochemical (IHC) staining of indicated KPC1 PDAC (PIP, PIL, PILiver) and KP1 LUAD (LIL, LIP, LILiver) derived-tumors treated with vehicle (V) or combined trametinib (1 mg/kg) and palbociclib (100 mg/kg) (T/P) for 2 weeks. Quantification of the percentage of SA-β-gal+ area and the number of Ki67+ and pRb+ cells per field are shown inset (n = 2-4 independent tumors per group). Scale bar, 50μm. c, Immunofluorescence staining of indicated KPC1 PDAC (PIP, PIL, PILiver) and KP1 LUAD (LIL, LIP, LILiver) derived-tumors treated as in (b). Quantification of the percentage of GFP+ (green) tumor cells expressing p21 (cyan) is shown inset (n = 2-4 independent tumors per group). Scale bar, 50μm. d, GFP+ tumor cells were FACS sorted from indicated tumors and extracted RNA subjected to RNA-seq analysis (n = 2-4 independent samples per group). Gene Set Enrichment Analysis (GSEA) of RNA-seq data using an established senescence gene set is shown. NES, normalized enrichment score. P values in d were calculated using two-sided, Kolmogorov-Smirnov test. Error bars, mean ± SEM. IHC experiments were repeated at least twice and representative images are shown.
Extended Data Fig. 2 Suppression of NK immunity specific to pancreas TME following T/P-induced senescence.
a-b, KPC2 PDAC or KP2 LUAD tumor cells expressing GFP were injected i.v. or orthotopically into the pancreas of 8-12 week old C57BL/6 female mice. Following tumor formation, mice were treated with vehicle (V) or combined trametinib (1 mg/kg) and palbociclib (100 mg/kg) (T/P) for 2 weeks. Flow cytometry analysis of NK cell numbers and degranulation in PDAC (PIP, PIL) (a) and LUAD-derived tumors (LIL, LIP) (b) are shown (PIP V, n = 3; PIP T/P, n = 4; PIL V and PIL T/P, n = 8; LIL V, n = 6; LIL T/P, n = 7; LIP V and T/P, n = 9 mice). c, Flow cytometry analysis of NK cell numbers and degranulation in spleens of mice with KPC1-derived PIP tumors treated as in (a) (n = 5 mice per group). d, Kaplan-Meier survival curve of mice with KPC2-derived PIP tumors treated with vehicle, combined trametinib (1 mg/kg) and palbociclib (100 mg/kg), and/or depleting antibodies against NK1.1 (PK136; 250 μg) or CD8 (2.43; 200 μg) (V, n = 5; T/P, T/P + αNK1.1 and T/P + αCD8, n = 8 mice). e, IVIS images showing luciferase signaling in KPC1-derived PIL tumors following treatment as in (a). Right, quantification of total luminescence in the thoracic region (V, n = 5; T/P and T/P + αNK1.1, n = 8 mice). f, Waterfall plot of the response of KPC1-derived PIP tumors following 2 week treatment with vehicle, combined trametinib (1 mg/kg) and palbociclib (100 mg/kg), and/or an NK1.1 depleting antibody (PK136; 250 μg) (V and T/P, n = 5; T/P + αNK1.1, n = 6 mice). g, Waterfall plot of the response of KPC2-derived PIP tumors following 2 week treatment with vehicle, combined trametinib (1 mg/kg) and palbociclib (100 mg/kg), and/or an NK1.1 (PK136; 250 μg) or CD8 (2.43; 200 μg) depleting antibody (V, n = 5; T/P, n = 7; T/P + αNK1.1 and T/P + αCD8, n = 8 mice). h, Kaplan-Meier survival curve of mice with KPC1-derived PIL tumors treated with vehicle, combined trametinib (1 mg/kg) and palbociclib (100 mg/kg), and/or depleting antibodies against CD8 (2.43; 200 μg) or CD4 (GK1.5; 200 μg) (V, n = 5; T/P, T/P + αCD4 and T/P + αCD8, n = 7 mice). i, Flow cytometry analysis of CD4+ and CD8+ T cell numbers and degranulation in KPC1 PDAC (PIP, PIL, PILiver) and KP1 LUAD-derived tumors (LIL, LIP, LILiver) grown in different organs and treated as in (a) (PIP V and T/P, n = 5 or 10; PIL V, n = 5 or 9; PIL T/P, n = 5 or 10; LIL V, n = 3; LIL T/P, n = 5; LIP V, n = 5 or 13; LIP T/P, n = 6 or 15; PILiver V, n = 9; PILiver T/P, n = 10; LILiver V, n = 8; LILiver T/P, n = 10 mice). Data represents pool of 3 independent experiments. P values in a-c, e-g, and i were calculated using two-tailed, unpaired Student’s t-test, and those in d and h were calculated using log-rank test. Error bars, mean ± SEM.
Extended Data Fig. 3 Repression of pro-inflammatory SASP gene expression specific to the pancreas TME following T/P treatment.
a, Heatmaps showing fold change in IFNα (left), IL-12 (middle), and TNFα pathway genes (right) following T/P treatment in indicated tumor settings from RNA-seq data in Fig. 2a (n = 2-4 independent samples per group). b, IHC staining of indicated KPC1 PDAC (PIP, PIL) and KP1 LUAD (LIL, LIP) derived-tumors grown in different organs and treated with vehicle (V) or combined trametinib (1 mg/kg) and palbociclib (100 mg/kg) (T/P) for 2 weeks. H-score quantification of CCL2 and CXCL10 staining intensity is shown inset (n = 2-3 independent tumors per group). Scale bars, 50μm. Error bars, mean + SEM. c, Transcription factor enrichment analysis showing transcriptional regulators of targets differentially expressed in tumors in the lungs (LIL, PIL) following T/P treatment. IHC experiments were repeated at least twice and representative images are shown.
Extended Data Fig. 4 Tumors in the pancreas TME are enriched for H3K27me3 repressive chromatin marks at SASP gene loci.
a, Heatmaps of normalized genome-wide H3K27me3 signaling intensities of consensus peaks from CUT&Tag analysis of KPC1 cells treated with vehicle or trametinib (25 nM) and palbociclib (500 nM) in vitro for 8 days, or KPC1 cells FACS sorted from transplanted PDAC tumors in C57BL/6 female mice treated with vehicle or trametinib (1 mg/kg) and palbociclib (100 mg/kg) for 2 weeks (n = 2-4 independent samples per group). b, Normalized H3K27me3 peak intensities of 87 SASP genes (see Table 47) from CUT&Tag analysis samples in (a) (n = 2-4 per independent samples group). c, Genome browser tracks showing H3K27me3 occupancy at pro-inflammatory SASP gene loci from CUT&Tag analysis samples in (a) (n = 2-4 independent samples per group).
Extended Data Fig. 5 PDAC and LUAD tumor cells have a similar pro-inflammatory SASP response to T/P-induced senescence in vitro.
a, Cytokine array analysis of pro-inflammatory SASP factors in murine PDAC and LUAD cell lines treated with vehicle or combined trametinib (25 nM) and palbociclib (500 nM) for 8 days (n = 2 independent samples per group). #, outside the detectable limit. b, Normalized expression levels of pro-inflammatory SASP genes in human PDAC and LUAD cell lines following treatment as in (a) from analysis of RNA-seq data generated in Ruscetti et al. 201815 (n = 2 independent samples per group).
Extended Data Fig. 6 Suppression of EZH2-mediated H3K27me3 induces pro-inflammatory SASP and immunomodulatory cell surface molecules following T/P treatment in PDAC cells.
a, Representative clonogenic assay images (from 3 biological replicates) of KPC1 cells harboring shRen or shEzh2 shRNAs replated in the absence of drugs after 8-day pre-treatment with vehicle or combined trametinib (25 nM) and palbociclib (500 nM) (top). Bottom, representative SA-β-gal staining (from 3 biological replicates) of KPC1 cells harboring shRen or shEzh2 shRNAs and treated as above for 8 days. Scale bar, 50μm. Experiments were repeated at least twice with similar results. b, RT-qPCR analysis of senescence and SASP gene expression in KPC1 cells harboring shRen or shEzh2 shRNAs treated as in (a) (n = 3 independent samples per group). A.U., arbitrary units. c, Representative histograms (top) and quantification of mean fluorescent intensity (MFI) of MHC-I (H-2kb) expression (bottom) on KPC1 cells harboring shRen or shEzh2 shRNAs (left) (n = 3 independent samples per group) or parental KPC1 cells (right) treated with vehicle, combined trametinib (25 nM) and palbociclib (500 nM), and/or tazemetostat (5 μM) for 8 days (n = 6 independent samples per group). Experiments were repeated at least twice with similar results. d, Representative clonogenic assay images (from 3 biological replicates) of KPC1 cells replated in the absence of drugs after 8-day pre-treatment with vehicle, combined trametinib (25 nM) and palbociclib (500 nM), and/or tazemetostat (5 μM) (top). Bottom, representative SA-β-gal staining (from 3 biological replicates) of KPC1 cells treated as above for 8 days. Scale bar, 50μm. Experiments were repeated at least twice with similar results. e, RT-qPCR analysis of SASP gene expression in human PANC-1 PDAC cells treated with vehicle, trametinib (25 nM), palbociclib (500 nM), and/or GSK126 (1 μM) for 8 days (n = 3 independent samples per group). A.U., arbitrary units. f, Heatmaps of normalized genome-wide H3K27me3 signaling intensities from CUT&Tag analysis of KPC1 cells harboring Ren or Ezh2 shRNAs treated with vehicle or trametinib (25 nM) and palbociclib (500 nM) for 8 days (n = 2-4 independent samples per group). g, Genome browser tracks showing H3K27me3 occupancy at pro-angiogenic SASP gene loci. P values in b, c, and e were calculated using two-tailed, unpaired Student’s t-test. Error bars, mean ± SEM.
Extended Data Fig. 7 EZH2 knockdown in the KPC2 PDAC orthotopic transplant model potentiates anti-tumor NK and CD8+ T cell immunity and long-term tumor regressions following T/P treatment.
a, Ultrasound quantification of initial PDAC tumor volume 1-week post-transplantation of KPC1 or KPC2 cells harboring shRen or shEzh2 shRNAs into 8-12 week old C57BL/6 female mice prior to enrollment in treatment cohorts (KPC1 shRen, n = 18; KPC1 shEzh2, n = 75; KPC2 shRen, n = 21; KPC2 shEzh2, n = 20 mice). Data represents pool of 6 independent experiments. b, Western blots of shRen or shEzh2 KPC1 orthotopic PDAC tumors treated with vehicle or trametinib (1 mg/kg) and palbociclib (100 mg/kg) for 2 weeks. c, IHC staining of KPC1 and KPC2 orthotopic PDAC tumors harboring shRen or shEzh2 shRNAs treated as in (b). H-score quantification of EZH2 expression is shown inset (n = 2-3 independent tumors per group). Scale bars, 50μm. d-e, Flow cytometry analysis of NK (d) and T cell (e) numbers and activation markers in KPC2 orthotopic PDAC tumors harboring indicated shRNAs treated as in (b) (shRen V, n = 6; shRen T/P, n = 7; shEzh2 V, n = 5; shEzh2 T/P, n = 7 mice). f, Flow cytometry analysis of F4/80+ macrophages in KPC1 orthotopic PDAC tumors harboring indicated shRNAs treated as in (b) (shRen V, n = 9; shRen T/P, n = 8; shEzh2 V, n = 6; shEzh2 T/P, n = 9 mice). Data represents pool of 2 independent experiments. g, Waterfall plot of the response of KPC2 orthotopic PDAC tumors harboring indicated shRNAs to treatment as in (b) (shRen V, n = 9; shRen T/P, n = 12; shEzh2 V, n = 7; shEzh2 T/P, n = 13 mice). Data represents pool of 2 independent experiments. h, Kaplan-Meier survival curve of mice with shEzh2 KPC2 orthotopic PDAC tumors treated with vehicle, combined trametinib (1 mg/kg) and palbociclib (100 mg/kg), and/or depleting antibodies against NK1.1 (PK136; 250 μg) or CD8 (2.43; 200 μg) (V, n = 5; T/P, T/P + αNK1.1, and T/P + αCD8, n = 8 mice). Dotted line indicates timepoint when mice were taken off of treatment. P values in a were calculated using One-way ANOVA followed by Tukey’s multiple comparison test, d-g using two-tailed, unpaired Student’s t-test, and h using log-rank test. Error bars, mean ± SEM. IHC staining and western blots were repeated at least twice and representative images are shown.
Extended Data Fig. 8 Combined EZH2 knockdown and T/P treatment reduces pancreatic metastasis growth and enhances NK and T cell immune surveillance in the lung.
a, KPC1 PDAC cells harboring shRen or shEzh2 shRNAs were injected i.v. into 8-12 week old C57BL/6 female mice. Following tumor formation in the lungs, mice were treated with vehicle (V) or combined trametinib (1 mg/kg body weight) and palbociclib (100 mg/kg body weight) (T/P) for 2 weeks. Quantification of lung tumor burden after 2 weeks of treatment is shown (shRen V and T/P, n = 4; shEzh2 V, n = 3; shEzh2 T/P, n = 5 mice). b-h, Flow cytometry analysis of total CD45+ immune cells (b), F4/80+ macrophages (c), NK cells (d), GZMB+ NK cells (e), CD4+ T cells (f), CD8+ T cells (g), and GZMB+ CD8+ T cells (h) in shRen or shEzh2 KPC1 PDAC tumors in the lung following treatment as in (a) (shRen V, n = 6; shRen T/P, n = 7; shEzh2 V, n = 8; shEzh2 T/P, n = 9 mice). P values in a-h were calculated using two-tailed, unpaired Student’s t-test. Error bars, mean ± SEM.
Extended Data Fig. 9 EZH2 blockade reduces T/P-induced blood vessel formation and promotes CCL2 and CXCL9/10 secretion that increases NK and CD8+ T cell infiltration into PDAC.
a, IHC staining of KPC1 orthotopic PDAC tumors harboring shRen or shEzh2 shRNAs from mice treated with vehicle or combined trametinib (1 mg/kg) and palbociclib (100 mg/kg) (T/P) for 2 weeks. Quantification of blood vessels per field are shown in the inset (n = 2-4 independent tumors per group). Scale bar, 50μm. b-c, Flow cytometry analysis of NK cell activation markers (b) and CD4+ and CD8+ T cell numbers (c) in KPC1 orthotopic PDAC tumors expressing control Empty or Ccl2 vectors and treated as in (a) (Empty V, n = 2 or 4; Empty T/P, n = 4 or 9; Ccl2 O/E V, n = 6 or 11; Ccl2 O/E T/P, n = 7 or 12 mice). Data represents pool of 3 independent experiments d, Flow cytometry analysis of CD4+ and CD8+ T cell numbers in shEzh2 KPC1 orthotopic PDAC tumors following treatment with vehicle, combined trametinib (1 mg/kg) and palbociclib (100 mg/kg), and/or a CCL2 depleting antibody (2H5; 200 μg) for 2 weeks (V and αCCL2, n = 3; T/P, n = 7; T/P + αCCL2, n = 9 mice). Data represents pool of 2 independent experiments. e, Flow cytometry analysis of NK cell numbers in shEzh2 KPC1 orthotopic PDAC tumors following treatment with vehicle, combined trametinib (1 mg/kg) and palbociclib (100 mg/kg), and/or a CXCR3 depleting antibody (CXCR3−173; 200 μg) for 2 weeks (V and T/P, n = 8; αCXCR3, n = 5; T/P + αCXCR3, n = 12 mice). Data represents pool of 2 independent experiments. P values in a-e were calculated using two-tailed, unpaired Student’s t-test. Error bars, mean ± SEM. IHC experiments were repeated at least twice and representative images are shown.
Supplementary information
Supplementary Information
Supplementary Fig. 1.
Supplementary Table 1
Supplementary Tables 1–49.
Source data
Source Data Figs. 1–8 and Extended Data Fig. 1–9
Statistical Source Data for Figs. 1–8 and Extended Data Figs. 1–9.
Source Data Figs. 4, 7 and Extended Data Fig. 7
Unprocessed western blots for Figs. 4 and 7 and Extended Data Fig. 7.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Chibaya, L., Murphy, K.C., DeMarco, K.D. et al. EZH2 inhibition remodels the inflammatory senescence-associated secretory phenotype to potentiate pancreatic cancer immune surveillance. Nat Cancer 4, 872–892 (2023). https://doi.org/10.1038/s43018-023-00553-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s43018-023-00553-8
