
Abstract
Enhanced glycolysis and accumulation of lactate is a common feature in various types of cancer. Intracellular lactate drives a recently described type of posttranslational modification, lysine lactylation (Kla), on core histones. However, the impact of lactylation on biological processes of tumour cells remains largely unknown. Here we show a global lactylome profiling on a prospectively collected hepatitis B virus-related hepatocellular carcinoma (HCC) cohort. Integrative lactylome and proteome analysis of the tumours and adjacent livers identifies 9,275 Kla sites, with 9,256 sites on non-histone proteins, indicating that Kla is a prevalent modification beyond histone proteins and transcriptional regulation. Notably, Kla preferentially affects enzymes involved in metabolic pathways, including the tricarboxylic acid cycle, and carbohydrate, amino acid, fatty acid and nucleotide metabolism. We further verify that lactylation at K28 inhibits the function of adenylate kinase 2, facilitating the proliferation and metastasis of HCC cells. Our study therefore reveals that Kla plays an important role in regulating cellular metabolism and may contribute to HCC progression.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others
Data availability
The processed data of this research, including genome, transcriptome, proteome and lactylome data, and their corresponding raw data, are all publicly available in the NODE database (The National Omics Data Encyclopedia) with accession code OEP002852 (https://www.biosino.org/node/project/detail/OEP002852). The CPTAC-HCC proteome data are available at the Proteomic Data Commons (PDC, https://pdc.cancer.gov/pdc/; accession node: PDC000198). Gene sets are available in Msigdb database (http://www.gsea-msigdb.org/gsea/msigdb/; C2 CP collection). Source data are provided with this paper.
References
Luo, J., Su, F., Chen, D., Shiloh, A. & Gu, W. Deacetylation of p53 modulates its effect on cell growth and apoptosis. Nature 408, 377–381 (2000).
Yu, J. et al. Histone lactylation drives oncogenesis by facilitating m(6)A reader protein YTHDF2 expression in ocular melanoma. Genome Biol. 22, 85 (2021).
Swamy, M. et al. Glucose and glutamine fuel protein O-GlcNAcylation to control T cell self-renewal and malignancy. Nat. Immunol. 17, 712–720 (2016).
Lin, R. et al. Acetylation stabilizes ATP-citrate lyase to promote lipid biosynthesis and tumor growth. Mol. Cell 51, 506–518 (2013).
Zhang, D. et al. Metabolic regulation of gene expression by histone lactylation. Nature 574, 575–580 (2019).
Liberti, M. V. & Locasale, J. W. The Warburg Effect: how does it benefit cancer cells? Trends Biochem. Sci. 41, 211–218 (2016).
Sung, H. et al. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 71, 209–249 (2021).
Jiang, Y. et al. Proteomics identifies new therapeutic targets of early-stage hepatocellular carcinoma. Nature 567, 257–261 (2019).
Gao, Q. et al. Integrated proteogenomic characterization of HBV-related hepatocellular carcinoma. Cell 179, 561–577 e522 (2019).
Zhao, S. et al. Regulation of cellular metabolism by protein lysine acetylation. Science 327, 1000–1004 (2010).
Wang, Q. et al. Acetylation of metabolic enzymes coordinates carbon source utilization and metabolic flux. Science 327, 1004–1007 (2010).
Aggeler, R. et al. A functionally active human F1F0 ATPase can be purified by immunocapture from heart tissue and fibroblast cell lines. Subunit structure and activity studies. J. Biol. Chem. 277, 33906–33912 (2002).
Gregory, R. I. et al. The Microprocessor complex mediates the genesis of microRNAs. Nature 432, 235–240 (2004).
Krynetski, E. Y., Krynetskaia, N. F., Bianchi, M. E. & Evans, W. E. A nuclear protein complex containing high mobility group proteins B1 and B2, heat shock cognate protein 70, ERp60, and glyceraldehyde-3-phosphate dehydrogenase is involved in the cytotoxic response to DNA modified by incorporation of anticancer nucleoside analogues. Cancer Res. 63, 100–106 (2003).
Hanahan, D. & Weinberg, R. A. Hallmarks of cancer: the next generation. Cell 144, 646–674 (2011).
Moreno-Yruela, C. et al. Class I histone deacetylases (HDAC1-3) are histone lysine delactylases. Sci. Adv. 8, eabi6696 (2022).
Dong, L. Q. et al. Heterogeneous immunogenomic features and distinct escape mechanisms in multifocal hepatocellular carcinoma. J. Hepatol. 72, 896–908 (2020).
Scheuer, P. J. Classification of chronic viral hepatitis: a need for reassessment. J. Hepatol. 13, 372–374 (1991).
Finn, R. S. et al. Atezolizumab plus bevacizumab in unresectable hepatocellular carcinoma. N. Engl. J. Med. 382, 1894–1905 (2020).
Sondka, Z. et al. The COSMIC Cancer Gene Census: describing genetic dysfunction across all human cancers. Nat. Rev. Cancer 18, 696–705 (2018).
Yu, W., Dittenhafer-Reed, K. E. & Denu, J. M. SIRT3 protein deacetylates isocitrate dehydrogenase 2 (IDH2) and regulates mitochondrial redox status. J. Biol. Chem. 287, 14078–14086 (2012).
Ericksen, R. E. et al. Loss of BCAA catabolism during carcinogenesis enhances mTORC1 activity and promotes tumor development and progression. Cell Metab. 29, 1151–1165 e1156 (2019).
Wu, T. et al. Discovery of a carbamoyl phosphate synthetase 1-deficient HCC subtype with therapeutic potential through integrative genomic and experimental analysis. Hepatology 74, 3249–3268 (2021).
Maddocks, O. D. et al. Serine starvation induces stress and p53-dependent metabolic remodelling in cancer cells. Nature 493, 542–546 (2013).
Maddocks, O. D. & Vousden, K. H. Metabolic regulation by p53. J. Mol. Med (Berl.) 89, 237–245 (2011).
Zucman-Rossi, J., Villanueva, A., Nault, J. C. & Llovet, J. M. Genetic landscape and biomarkers of hepatocellular carcinoma. Gastroenterology 149, 1226–1239 e1224 (2015).
Senni, N. et al. β-catenin-activated hepatocellular carcinomas are addicted to fatty acids. Gut 68, 322–334 (2019).
Adebayo Michael, A. O. et al. Inhibiting glutamine-dependent mTORC1 activation ameliorates liver cancers driven by β-catenin mutations. Cell Metab. 29, 1135–1150.e1136 (2019).
ICGC/TCGA Pan-Cancer Analysis of Whole Genomes Consortium Pan-cancer analysis of whole genomes. Nature 578, 82–93 (2020).
Kim, H. et al. The DUSP26 phosphatase activator adenylate kinase 2 regulates FADD phosphorylation and cell growth. Nat. Commun. 5, 3351 (2014).
Chen, R. P. et al. Adenylate kinase 2 (AK2) promotes cell proliferation in insect development. BMC Mol. Biol. 13, 31 (2012).
Cai, F. et al. AK2 promotes the migration and invasion of lung adenocarcinoma by activating TGF-beta/Smad pathway in vitro and in vivo. Front. Pharm. 12, 714365 (2021).
van Horssen, R. et al. Modulation of cell motility by spatial repositioning of enzymatic ATP/ADP exchange capacity. J. Biol. Chem. 284, 1620–1627 (2009).
Lee, H. J. et al. AK2 activates a novel apoptotic pathway through formation of a complex with FADD and caspase-10. Nat. Cell Biol. 9, 1303–1310 (2007).
Gao, Q. et al. CXCR6 upregulation contributes to a proinflammatory tumor microenvironment that drives metastasis and poor patient outcomes in hepatocellular carcinoma. Cancer Res. 72, 3546–3556 (2012).
Prianichnikov, N. et al. MaxQuant software for ion mobility enhanced shotgun proteomics. Mol. Cell Proteom. 19, 1058–1069 (2020).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
DePristo, M. A. et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 43, 491–498 (2011).
Cibulskis, K. et al. Sensitive detection of somatic point mutations in impure and heterogeneous cancer samples. Nat. Biotechnol. 31, 213–219 (2013).
Koboldt, D. C. et al. VarScan: variant detection in massively parallel sequencing of individual and pooled samples. Bioinformatics 25, 2283–2285 (2009).
Karczewski, K. J. et al. The ExAC browser: displaying reference data information from over 60 000 exomes. Nucleic Acids Res. 45, D840–D845 (2017).
Genomes Project, C. et al. A global reference for human genetic variation. Nature 526, 68–74 (2015).
Sherry, S. T. et al. dbSNP: the NCBI database of genetic variation. Nucleic Acids Res. 29, 308–311 (2001).
Wang, K., Li, M. & Hakonarson, H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 38, e164 (2010).
Cingolani, P. et al. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly. (Austin) 6, 80–92 (2012).
Kim, D. et al. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 14, R36 (2013).
Newman, A. M. et al. FACTERA: a practical method for the discovery of genomic rearrangements at breakpoint resolution. Bioinformatics 30, 3390–3393 (2014).
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
Robinson, M. D., McCarthy, D. J. & Smyth, G. K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
Xu, J. et al. In-depth profiling and quantification of the lysine acetylome in hepatocellular carcinoma with a trapped ion mobility mass spectrometer. Mol. Cell Proteom. 21, 100255 (2022).
Chai, X. et al. Quantitative acetylome analysis reveals histone modifications that may predict prognosis in hepatitis B-related hepatocellular carcinoma. Clin. Transl. Med 11, e313 (2021).
Satpathy, S. et al. A proteogenomic portrait of lung squamous cell carcinoma. Cell 184, 4348–4371.e4340 (2021).
Colaert, N., Helsens, K., Martens, L., Vandekerckhove, J. & Gevaert, K. Improved visualization of protein consensus sequences by iceLogo. Nat. Methods 6, 786–787 (2009).
Ashburner, M. et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 25, 25–29 (2000).
Mi, H. et al. PANTHER version 16: a revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 49, D394–D403 (2021).
Ruepp, A. et al. CORUM: the comprehensive resource of mammalian protein complexes–2009. Nucleic Acids Res. 38, D497–D501 (2010).
Shannon, P. et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504 (2003).
Kanehisa, M. & Goto, S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30 (2000).
Liberzon, A. et al. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 1, 417–425 (2015).
Barbie, D. A. et al. Systematic RNA interference reveals that oncogenic KRAS-driven cancers require TBK1. Nature 462, 108–112 (2009).
Acknowledgements
We acknowledge the assistance of Y. Weng (PTM BIO) for developing and evaluating method of mass spectrometry, L. Dong (Zhongshan Hospital, Fudan University) for the construction of TMAs and J. Shi (Zhongshan Hospital, Fudan University) for the supply of materials in multiplexed immunohistochemistry. This work was supported by the National Natural Science Foundation of China (grant Nos. 81961128025 to Q.G., 81973164 to H.H. and 81872321 to J.S.) and by Research Projects from the Science and Technology Commission of Shanghai Municipality (grant Nos. 19XD1420700 to Q.G., 21JC1401200 to Q.G. and 21JC1410100 to J.F.), Shanghai Municipal Key Clinical Specialty and Shanghai Municipal Science and Technology Major Project.
Author information
Authors and Affiliations
Contributions
Q.G., H.H. and J.F. conceived and designed the research. Z.Y., J.M., S.Z. and S.Q. coordinated acquisition, distribution and quality evaluation of HCC tumour, and adjacent liver tissue samples. H.H., Z.Y., C.Y., X.R. and X.S. directed and performed analyses and quality control of mass spectrometry data. Z.Y., X.R., H.H., S.C. and Y.W. analysed genome and transcriptome data. J.M. and Z.Y. performed the tissue microarray immunohistochemistry experiments. C.Y., P.P., X.R. and Z.Y. performed HCC cell line experiments. Q.G., J.F., X.W. and J.Z. interpreted data in context of HCC biology and provided clinical guidance. Z.Y., C.Y. and X.R. wrote the manuscript. Q.G., H.H. and J.F. revised the manuscript and supervised the study.
Corresponding authors
Ethics declarations
Competing interests
S.C. is an employee of Burning Rock Biotech. The other authors declare no competing interests.
Peer review
Peer review information
Nature Metabolism thanks Sabine Colnot and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Alfredo Giménez-Cassina, in collaboration with the Nature Metabolism team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Overview of the research pipeline and quality control for the mass spectrometry platform.
a, Schematic description of research pipeline. b, Illustration of 52 cases of HCC patients included in the individual omics profiling. The genome, transcriptome, proteome and lactylome experiments are shown, respectively. The main clinical information of each patient is shown. Tumor2: Two tumour samples are obtained from each patient with multifocal HCC (n = 6). c, Distribution of score and mass error of differently charged peptides. d, Spearman correlation coefficients calculated from the proteome (left) and lactylome (right) of each biological replicate used for quality control. e, The number of identified Kla sites from all 110 samples from 52 HCC patients.
Extended Data Fig. 2 Basic properties of Kla in HCC and lactylprotein complex.
a, Distribution of tumour/adjacent ratio of Kla sites. The displayed Kla sites all satisfy p < 0.05 in the comparison between the tumours and adjacent livers. b, Intersection of Kla and Kac sites from five cohorts (all sites, left; sites on metabolism proteins, right). c, Number of Kla sites per protein in the tumours and adjacent livers. d, Subcellular location of all lactylproteins in the tumours and adjacent livers. e, Protein complexes enriched in the lactylproteins differentially lactylated between tumours and adjacent livers. The size of nodes represents the number of Kla sites identified in each protein subunit. The edge represents the interaction between one subunit and the other or itself recorded in the CORUM database. f, Number of detected Kla sites on different histones. N = 58 and 52 for tumour and adjacent liver samples. g, Number of detected histone Kla sites in tumour or adjacent normal tissues. ****P < 0.0001. h, Kla intensity on different histones in tumour or adjacent normal tissues. Tumour versus Adjacent: ***P = 0.00014, ***P = 0.00015. i, Differentially lactylated Kla sites on histones detected. Tumour versus Adjacent: *P = 0.02, ****P < 0.0001, ***P = 0.00054, ***P = 0.00037, ***P = 0.00054, *P = 0.027, **P = 0.0011, **P = 0.013. Box plot centres represent the median and lower and upper hinges represent the first and third quartiles (25th and 75th percentiles), the whiskers extend to the largest and smallest value (g–i). P value is calculated by two-sided Wilcoxon rank-sum test (g–i).
Extended Data Fig. 3 Cellular Kla can be affected by multiple factors.
a, Protein expression of GLUT1/SLC2A1, MCT1/SLC16A1, MCT4/SLC16A3 in Fudan-Kla and CPTAC-HCC cohort. Tumour versus Adjacent (Fudan-Kla cohort): *P = 0.019, ****P < 0.014; Tumour versus Adjacent (CPTAC-HCC cohort): *P = 0.048, ****P < 0.0001. b, SLC2A1, SLC16A1 and SLC16A3 were knocked down using shRNA in HepG2 cells, respectively. Cellular Kla intensity was inspected by immunoblots. c, Protein expression of indicated genes in tumour and adjacent tissue derived from Fudan-Kla cohort and CPTAC-HCC cohort. Tumour versus Adjacent (Fudan-Kla cohort): ****P < 0.0001, ****P < 0.0001, ****P < 0.0001, ****P < 0.0001. Tumour versus Adjacent (CPTAC-HCC cohort): ****P < 0.0001, ****P < 0.0001, ****P < 0.0001, ****P < 0.0001, **P = 0.0091, ***P = 0.00015, ****P < 0.0001, ****P < 0.0001, ****P < 0.0001, *P = 0.043. d, Spearman correlation coefficients between Kla intensity and expression of different proteins. x axis represents the protein expression of EP300, y axis represents the protein expression of indicated erasers. Number at four corners represents the number of Kla sites on metabolism/non-metabolism proteins. e, HepG2 cells were treated with 10 μM A485 for 48 h, followed by Kla detection. f, HepG2 cells were transfected with shRNA-HDAC1, shRNA-HDAC2 and shRNA-HDAC3, followed by Kla detection. N = 58 and 52 for tumour and adjacent liver samples (Fudan-Kla cohort), N = 159 and 159 for tumour and adjacent liver samples (CPTAC-HCC cohort) (a,c). Box plot centres represent the median and lower and upper hinges represent the first and third quartiles (25th and 75th percentiles), the whiskers extend to the largest and smallest value (a,c). P value is calculated by two-sided Wilcoxon rank-sum test (a,c). Experiments were performed three times (b,e,f).
Extended Data Fig. 4 Pathway overrepresentation of Kla sites in each quadrant.
a, Pathway enrichment of lactylproteins. Pathways with adjusted P value < 0.01 are shown. All pathways are composed of upregulated Kla in tumours compared with adjacent livers. There are no pathways for downregulated ones in the tumours compared with the adjacent livers that meet the need of adjusted P value < 0.01. b, Barplots showing the highly represented pathways with adjusted P value < 0.01. The quadrants marked on top correspond to Fig. 2c, representing the number of Kla sites on differentially expressed Oncogene/TSG between tumours and adjacent livers. c, Overlap among mRNA, protein, lactylprotein for the Cancer Gene Census (CGC). The schematic diagram of the quadrants in lactylprotein corresponds to the quadrants in Fig. 2a. P value is calculated by hypergeometric test and adjusted by Benjamini–Hochberg method (a,b).
Extended Data Fig. 5 Lactylation of AK2-K28 is associated with unfavourable prognosis in HCC.
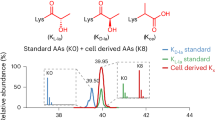
a, Transcriptomic level of AK2 at the pan-cancer level. Data is derived from PCAWG database. N = 187, 533, 139, 79, 265, 183, 550, 91, 196, 323, 168, 1218, 450, 553, 122, 572, 87, 555, 45, 80, 171, 566, 427, 57, 576, 606, 492, 424, 309, 474, 48 and 310 for PCPG, LGG, TGCT, ACC, SARC, PAAD, PRAD, KICH, ESCA, KIRP, GBM, BRCA, STAD, LUSC, THYM, THCA, MESO, UCEC, CHOL, UVM, READ, HNSC, BLCA, UCS, LUAD, KIRC, COAD, LIHC, OV, SKCM, DLBC, CESC samples. b, Kaplan–Meier curve of overall survival for AK2 protein expression in tumours of CPTAC-HCC patients (AK2 + high, n = 80; GPC3 + panKla+AK2 + low, n = 79). c, Quantitative analysis of differential expression of AK2 between tumours and adjacent livers (left) and the western blot validation (right). N = 58 and 52 for tumour and adjacent liver samples. d, Quantitative analysis of differentially lactylated Kla sites on AK2 between tumours and adjacent livers. N = 58 and 52 for tumour and adjacent liver samples. e, The MS/MS spectra (left) and extracted ion chromatograms (right) of the synthetic AK2-K28la peptide (top) corresponding to the sequence of the in vivo peptide (AVLLGPPGAGKlaGTQAPR) (bottom). f, Associations of AK2 protein expression with the corresponding pathway score, clinicopathologic factors, and multiomics profiling. g, Associations of AK2-K28la with driver mutation, pathway scores, clinicopathologic factors and multiomics profiling. Box plot centres represent the median and lower and upper hinges represent the first and third quartiles (25th and 75th percentiles), the whiskers extend to the largest and smallest value (a,c,d). The P value is obtained from the two-sided Wilcoxon rank-sum test (c,d,f,g).
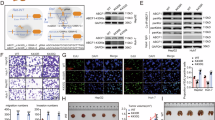
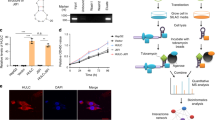
Extended Data Fig. 6 AK2-K28la weakens AK2 enzymatic activity and Kla of several important metabolic enzymes.
a, HepG2 and Hep3B cells ectopically expressing Flag-tagged AK2 were stimulated with 10 mM lactate for 24 h. Cell lysates were subjected to immunoprecipitation with Flag-M2 beads. Inputs and eluates were analysed by immunoblots. b, HepG2 ectopically expressing Flag-tagged AK2 were stimulated with 10 mM lactate for 24 h. Cell lysates were subjected to immunoprecipitation with Flag-M2 beads. Inputs and eluates were analysed by immunoblots. Time course of Kla of AK2 in HepG2 cells was revealed by immunoprecipitation. c, Kinetic measurements were performed for the activities of Flag-AK2-WT, K28R, K28Q, K28E and K28A mutants in HepG2 cells. d, Depletion and rescue efficiency of AK2 in HepG2 cells were detected by Western blot. e, Metabolome quantification of ADP and AMP in different groups indicated. The horizontal line represents the median. f, Lactylation of AK2 results in enhanced cell proliferation in Hep3B cells. Error bars represent ±s.d. for triplicate experiments. Data are presented as mean ± s.e.m.; *P < 0.05. g, Transwell assay on migration capability of AK2-lactylation. HepG2 cells transfected with Flag-AK2 with or without treatment of 10 mM Lactate for 24 h. After 48 h, cells that migrated to the inferior membrane were stained. h, Expression of genes showing statistical difference in the comparison of WT_nontreated and WT_treated (two-sided Student’s t test, P < 0.05) but not in K28R_nontreated and K28R_treated (two-sided Student’s t test, P ≥ 0.05). i, Gene enrichment analysis results for genes upregulated in the comparison of WT_nontreated and WT_treated but not in the comparison of K28R_nontreated and K28R_treated. j, Transcriptomic expression of Caspases8, 9, 3 and 7 in different groups of HepG2 cells as indicated. N = 3 biological independent samples for each group. k, Lactylation levels of HADH, ADH5, DLD and ALDH7A1 proteins are elevated upon 10 mM lactate treatment for 24 hours in the indicated cells. Mutation of Lys93 and Lys528 to Arg leads to remarkably reduced lactylation level of ALDH7A1 after treatment with 10 mM lactate. Experiments were performed three times (a,b,d,k). P value is calculated with two-sided Wilcoxon rank-sum test (e,j), two-tailed Student’s t test (f), or hypergeometric test and adjusted by Benjamini–Hochberg method (i). Box plot centres represent the median and lower and upper hinges represent the first and third quartiles (25th and 75th percentiles), the whiskers extend to the largest and smallest value (j).
Supplementary information
Supplementary Table 1
The liver samples used in this study were obtained from the Zhongshan Hospital, Fudan University in China. There is no bias in the selection of patients. Every patient offers one non-tumour adjacent tissue sample and one to two tumour tissue samples. See Methods for details.
Supplementary Table 2
Identification information of Kla sites.
Supplementary Table 3
Gene list of capture-based genome sequencing.
Supplementary Table 4
Sequence data of AK2-K28R mutant was shown.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Fig. 7
Statistical source data.
Source Data Fig. 8
Statistical source data.
Source Data Fig. 8
Unprocessed western blots.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 3
Unprocessed western blots.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 5
Unprocessed western blots.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 6
Unprocessed western blots.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Yang, Z., Yan, C., Ma, J. et al. Lactylome analysis suggests lactylation-dependent mechanisms of metabolic adaptation in hepatocellular carcinoma. Nat Metab 5, 61–79 (2023). https://doi.org/10.1038/s42255-022-00710-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s42255-022-00710-w
