
Abstract
Mesenchymal stromal cells (MSCs) tend to infiltrate into tumors and form a major component of the tumor microenvironment. Our previous work demonstrated that tumor necrosis factor α (TNFα)-activated MSCs significantly promoted tumor growth. However, the role of TNFα-treated MSCs in tumor metastasis remains elusive. Employing a lung metastasis model of murine breast cancer, we found that TNFα-activated MSCs strikingly enhanced tumor metastasis compared with normal MSCs. We analyzed the chemokine profiles and found that the expression of CCL5, CCR2 and CXCR2 ligands were enhanced in TNFα-activated MSCs. Using genetic or pharmacological strategies to inhibit CCL5 or CCR2, we demonstrated that CCL5 and CCR2 ligands were indispensable in supporting TNFα-activated MSCs to promote tumor metastasis. Analysis of immune cells revealed that CXCR2 ligands (CXCL1, CXCL 2 and CXCL5) expressed by TNFα-activated MSCs efficiently recruited CXCR2+ neutrophils into tumor. These neutrophils were responsible for the pro-metastatic effect of MSCs since inhibition of this chemotaxis abolished increased neutrophil recruitment and tumor metastasis. The interaction between neutrophils and tumor cells resulted in markedly elevated metastasis-related genes by tumor cells, including CXCR4, CXCR7, MMP12, MMP13, IL-6 and TGFβ. Importantly, in IL8high human breast cancer samples, we also observed similar alterations of gene expression. Collectively, our findings demonstrate that TNFα-activated MSCs promote tumor metastasis via CXCR2+ neutrophil recruitment.
Similar content being viewed by others

Introduction
Solid tumors contain many kinds of cells besides tumor cells per se, such as immune cells, epithelial cells and mesenchymal stromal cells (MSCs).1, 2, 3 They help in building the tumor microenvironment (TME). The precursors of MSCs are a group of fibroblast-like adult stem cells that mainly exists in the bone marrow.4 Under inflammatory conditions such as those caused by damaged tissues, MSCs rapidly migrate to sites of wounding.5, 6 As tumors have many characteristics of a wound that never heals,7 we and others found that MSCs can also be recruited to tumor sites and regulate tumor development in multiple ways.8, 9, 10, 11, 12 Among these, MSC-mediated regulation of immune cells has gained much more attention in recent years.13, 14, 15
MSCs have a close relationship with immune cells. Our previous work found that under strong inflammatory conditions, MSCs exhibited immunosuppressive characteristics.16, 17 MSCs recruit immune cells to their surroundings through secretion of large amounts of chemokines. MSCs then produce immunosuppressive factors, such as nitric oxide and indoleamine 2, 3-dioxygenase, to suppress immune cell function. These properties suggest that MSCs have a potential ability to treat immune disorders. However, we also found that under weak inflammatory conditions, MSCs can enhance immune responses, as pro-inflammatory cytokine levels were inadequate to elicit sufficient nitric oxide and indoleamine 2, 3-dioxygenase production by MSCs.18 Thus, MSCs have a potential role in regulating immune responses in the TME. As one example, our previous work has demonstrated that tumor-derived MSCs (T-MSCs) significantly enhanced tumor growth through the recruitment of macrophages.13 These T-MSCs expressed high levels of CCR2 ligands, which were responsible for macrophage accumulation within tumor. Tumor necrosis factor α (TNFα)-activated MSCs mimicked T-MSCs in their chemokine production profile and ability to promote tumor growth. However, whether MSCs influenced tumor metastatic potential in an immune response dependent way was not further studied.
Tumor metastasis is the major cause of human cancer death. Recent studies from animal experiments suggested that tumor-infiltrating immune cells contribute to the metastatic cascade.19, 20, 21 A substantial amount of clinical data also indicate that tumor infiltration of certain types of immune cells correlates with poor prognosis of patients with cancer.22, 23 Among these, tumor-associated neutrophils are an important candidate to influence tumor metastasis because of the large numbers of infiltrating tumor-associated neutrophils found in tumor tissue.24 There are a substantial number of studies that suggest that tumor-associated neutrophils are important in promoting tumor metastasis, possibly through secreting matrix metalloproteinases (MMPs) and other soluble molecules.25, 26 The underlying mechanism of the recruitment of neutrophils into tumors has not been fully studied. Chemokines are important for neutrophil recruitment. However, different types of tumors may use different chemokines as neutrophil chemoattractants, such as CXCL5 in breast cancer,27 CXCL6 in gastrointestinal tumor28 and HMGB1 in melanoma.29 Whether there are universal mechanisms across different tumor types needs to be further explored.
In this study, we employed a murine breast cancer tumor model and found TNFα-activated MSCs significantly promoted tumor metastasis. By secreting CXCR2 ligands, these TNFα-activated MSCs efficiently recruited neutrophils into tumors. Inhibition of this chemotaxis reversed the pro-metastatic effect of MSCs. A co-culture system showed evidence that neutrophils activated tumor cells to express higher levels of metastasis-related genes. Thus, our study identifies a novel MSC/neutrophil/tumor cell axis, which is critical in neutrophil recruitment and subsequent tumor metastasis in breast cancer.
Results
TNFα-activated MSCs promote tumor metastasis
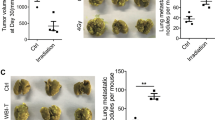
To study the effect of TNFα-activated MSCs on tumor metastasis, we co-injected 4T1 breast cancer cells with murine bone marrow-derived MSCs into the fat pad of the BALB/c mice. We found that TNFα-activated MSCs markedly promoted tumor metastasis to lung, whereas MSCs without cytokine treatment showed no effect (Figure 1a). Growth of the primary tumor was not altered by TNFα-activated MSCs (Supplementary Figures S1a and b). This result indicated that MSCs displayed greater potential to promote tumor metastasis when activated by TNFα. Animal survival analysis also showed that TNFα-activated MSCs markedly shortened the life span of tumor-bearing mice (Figure 1b).

TNFα-activated MSCs promote tumor metastasis. (a) 4T1 breast cancer cells (4 × 105) were co-injected with MSCs (1 × 105) into the fat pad of BALB/c mice; the control group received 4T1 cells alone. MSCs were activated by TNFα (10 ng/ml) for 24 h. Mice were euthanized after 4 weeks. Metastatic tumor nodules in the lung were counted. (b) Animal survival analysis. BALB/c mice received 4T1 cells alone or with different MSCs as described in a. (c) 4T1 breast cancer cells (4 × 105) were injected into fat pad of BALB/c mice. Mice were euthanized after 14 days. The concentration of TNFα in serum was tested by ELISA assay. TNFα levels in serum of normal mice served as a control. (d) Animal model was as described in a. Metastatic tumor nodules in the lung were counted. (e and f) For the wound-healing assay, cultured 4T1 cells were first starved for 24 h. Wound closures were photographed (e) and statistically analyzed (f) at 24 h after wounding (scale bars: 700 μm). Data are shown as means±s.e.m. Statistical significance was assessed by unpaired, two-tailed Student’s t-test.
The TME has a crucial role in modulating the properties of MSCs. Our previous study demonstrated that T-MSCs promoted tumor growth more potently than normal MSCs.13 TNFα-pretreated MSCs mimicked T-MSCs in promoting tumor growth. In our 4T1 breast cancer model, the concentration of TNFα in the serum was much higher in tumor-bearing mice compared with WT mice (Figure 1c). We therefore wondered if T-MSCs can exert a similar pro-metastatic effect as seen for TNFα-pretreated MSCs. We co-injected T-MSCs with 4T1 cells and as expected, lung metastasis were markedly increased (Figure 1d).
Our previous study showed that both T-MSCs and TNFα-activated MSCs exhibited altered patterns of gene expression in comparison with normal MSCs, especially in the levels of expression of chemokine and growth factor genes. There are also reports that suggest that MSCs promote tumor metastasis through directly enhancing tumor migration.10, 30 Based on these observations, we hypothesized that TNFα-activated MSCs facilitated tumor metastasis through directly secreting chemokines and growth factors. To test this, we examined the effect of MSCs in a wound-healing assay. Surprisingly, we found that cultured medium derived from TNFα-activated MSCs failed to enhance tumor cell migration as monitored by wound healing (Figures 1e and f). This suggests that TNFα-activated MSCs promote tumor metastasis through an indirect mechanism.
TNFα-activated MSCs mediated tumor metastasis promotion is independent of CCL5 and CCR2 ligands
We and others have demonstrated that MSCs can recruit immune cells and modify their functions through secretion of multiple chemokines and growth factors.16 Therefore, it is possible that MSCs promote metastasis indirectly by recruiting immune cells secondary to secretion of chemokines. To test this, we compared chemokine levels produced by different groups of MSCs using a qPCR array. Consistent with previous reports, multiple chemokines were upregulated in either T-MSCs or TNFα-activated MSCs (Figure 2a). Interestingly, among these genes, seven chemokines were significantly upregulated in both T-MSCs and TNFα-activated MSCs. These chemokines could be generally divided into three groups according to the chemokine receptor to which they bound, that is, CCR5, CCR2 and CXCR2, respectively (Figure 2a). Next we wanted to explore which group of chemokines was responsible for the pro-metastatic effect of MSCs.

TNFα-activated MSCs promote tumor metastasis did not through CCL5 and CCR2 ligands. (a) Data sets of chemokine mRNA expression by MSCs were obtained through qPCR array. Only the chemokines with more than fivefold changes are shown. (b and c) Animal experiments were as described in Figure 1a. CCL5-KO MSCs were activated by TNFα (10 ng/ml) for 24 h. Metastasis lung tumor nodules were identified by whole-lung images (stained by bouin’s solution), H&E staining (b) (scale bars: 500 μm) and counted (c). (d) BALB/c mice received 4T1 cells alone or with different MSCs as described in Figure 1a. Mice were euthanized after 10 days. Single-cell suspensions prepared from tumor tissues were analyzed for the frequency of CD11b+ F4/80+ macrophages by flow cytometry. (e and f) Animal experiments were as described in Figure 1a. RS504393 was injected intraperitoneally (2 mg/kg) every day. Tumors were excised after 4 weeks. Metastatic tumor nodules in the lung were detected by whole-lung images (stained by bouin’s solution) (e) and counted (f). Data are shown as means±s.e.m. Statistical significance was assessed by unpaired, two-tailed Student’s t-test.
Previous studies strongly suggested that CCL5 synthesized by MSCs can promote breast tumor metastasis in a human tumor cell xenograft model.10 Indeed, we observed higher levels of CCL5 in T-MSCs and TNFα-activated MSCs compared with untreated-MSCs, at both the mRNA and protein levels (Supplementary Figures 2a and b). This suggested that CCL5 might have a similar role in our tumor model. However, when we used CCL5-deficient MSCs, we found TNFα-activated CCL5-KO MSCs still promoted lung metastasis (Figures 2b and c), without affecting the primary tumor growth (Supplementary Figure 2c). Therefore, in our animal model, CCL5 is not the key factor promoting tumor metastasis.
The second chemokine group- CCL2, CCL7 and CCL8, share the same receptor, CCR2, a chemokine receptor that is essential for monocyte/macrophage trafficking, which is widely expressed on monocytes and macrophages. Our previous results showed that T-MSCs and TNFα-activated MSCs exerted their tumor-promoting effects by recruiting macrophages.13 To test whether these recruited macrophages also enhanced tumor metastasis, we first determined the number of macrophages in the tumors of either T-MSC or TNFα-activated MSC treated mice. Surprisingly, we observed no changes in either group, which was not consistent with our previous results (Figure 2d). A possible explanation was that, unlike the tumor cell lines we used in our previous study (B16 and EL4), the 4T1 tumor cells spontaneously expressed CCL2 (Supplementary Figure S3a). Therefore, 4T1 tumor cells may compensate for the function of MSCs by secreting CCR2 ligands.
Although the numbers of macrophages infiltrating in the tumors showed no differences, their properties may have changed. To exclude this possibility, we employed a CCR2 inhibitor, RS504393, to inhibit the activity of CCR2 ligands and the recruitment of macrophages. We found that primary tumor growth in all groups was significantly inhibited (Supplementary Figures S3b and c), indicating an important role for macrophages on tumor growth. When tumor metastases were studied, however, TNFα-activated MSCs still exerted similar pro-metastatic effects after RS504393 treatment (Figures 2e and f). Therefore, the pro-metastatic effect of MSCs was independent of CCR2 ligands.
Neutrophil accumulation is promoted by TNFα-activated MSCs
The third group of chemokines included CXCL1, CXCL2 and CXCL5, which all bind to CXCR2. Previous studies suggested that CXCR2 was abundant in neutrophils. Indeed, we found that, in the blood of tumor-bearing mice, almost all the CXCR2-positive immune cells were CD11b+Ly6G+ neutrophils, and nearly all the neutrophils expressed CXCR2 (Figure 3a). In contrast, circulating CD4+, CD8+ T cells, monocytes and macrophages expressed only very low to undetectable levels of CXCR2 (Figure 3b). Neither MSCs nor 4T1 tumor cells expressed CXCR2 (Supplementary Figure S4a). Therefore, neutrophils were clearly the primary if not the only TME-associated immune cell type that expressed high levels of CXCR2 in our 4T1 breast cancer system.

TNFα-activated MSCs recruit more neutrophils to tumor. (a and b) Peripheral blood cells from 4T1 tumor-bearing mice were stained for CD11b, Ly6G, CXCR2 and other immune cell markers and analyzed by flow cytometry. (c and d) BALB/c mice received 4T1 cells alone or with different MSCs as described in Figure 1a. Mice were euthanized after 10 days. Single-cell suspensions prepared from tumor tissues (c and d) or from peripheral blood (c) were analyzed for the frequency of CD11b+ Ly6G+ neutrophils by flow cytometry. Data are shown as means±s.e.m. Statistical significance was assessed by unpaired, two-tailed Student’s t-test.
We next examined the percentage of neutrophils in tumor-bearing mice. Interestingly, we found that mice injected with either T-MSCs or TNFα-activated MSCs showed evidence of increased intratumoral neutrophil infiltration (Figures 3c and d). The percentage of neutrophils in blood was also higher in these two groups (Figure 3c). Immunohistochemical analysis of primary tumor tissue also showed that TNFα-activation of MSCs resulted in increased neutrophils infiltration (Supplementary Figure S4b). These results clearly showed that T-MSCs and TNFα-activated MSCs increased neutrophil recruitment into the TME. The infiltration of other immune cells in the tumor was also examined. The percentage of CD4+, CD8+ T cells and monocytes showed no changes in mice injected with TNFα-activated MSCs compared with other groups (Supplementary Figures S4c, d and e), suggesting the specific accumulation of neutrophils.
TNFα-activated MSCs recruit neutrophils through CXCR2 ligands
As shown above, CXCR2 was abundantly expressed on neutrophils. Based on this and the fact that TNFα-activated MSCs secreted large amounts of CXCR2 ligands, we speculated that the increased infiltration of neutrophils within the TME was due to increased recruitment mediated by MSCs. Thus, we first compared the expression level of CXCR2 ligands in MSCs derived from different tissues of the tumor-bearing mice. Indeed, T-MSCs strongly expressed CXCR2 ligands (Figure 4a). However, in MSCs isolated from bone marrow and lung, these chemokines were only marginally detected. TNFα could activate bone marrow-MSCs to express CXCR2 chemokines in vitro at both mRNA (Figure 4b) and protein levels (Figure 4c). However, 4T1 tumor cells expressed a low level of CXCR2 ligands and did not respond to TNFα activation (Figure 4b). These data suggested that MSCs were likely to be the main source of CXCR2 chemokines in the tumor.

TNFα-activated MSCs secrete CXCR2 ligands and recruit neutrophils. (a) MSCs were generated from primary tumor, bone marrow and lung of the mice bearing 4T1 tumor. Expression of chemokines in MSCs was determined by qPCR at passage 1. (b) MSCs and 4T1 cells were cultured in vitro and activated by TNFα (10 ng/ml) for 12 h. Chemokine expression was determined by qPCR. (c) MSCs were cultured in vitro and activated by TNFα (10 ng/ml) for 24 h. Chemokine levels in the medium were determined by ELISA assay. (d) In the transwell assay, neutrophils were seeded in the top well and MSC-cultured medium (MSC-CM) placed in the bottom well. SB265610 was added to both the top and bottom wells. Data are shown as means±s.e.m. Statistical significance was assessed by unpaired, two-tailed Student’s t-test.
We next employed an in vitro transwell assay to determine whether CXCR2 ligands secreted by MSCs induced neutrophil chemotaxis. Neutrophils were freshly isolated from the blood of tumor-bearing mice (Supplementary Figure S5a). When the bottom chamber was filled with conditioned medium of TNFα-activated MSCs, we observed efficient migration of neutrophils (Figure 4d). A CXCR2-specific antagonist, SB265610, significantly inhibited neutrophil migration, suggesting that CXCR2 is required for neutrophil chemotaxis. Our results demonstrated that TNFα-activated MSCs attracted neutrophils through the secretion of CXCR2 ligands.
Neutrophils are responsible for the pro-metastatic effect of MSCs
To determine whether recruited neutrophils facilitate tumor metastasis, several experiments were performed. In the wound-healing assay, we found 4T1 cells migrated faster when co-cultured with neutrophils (Figures 5a and b). In animal experiments, we first determined whether co-injection of tumor cells with freshly isolated neutrophils could promote tumor metastasis. As shown in Figure 5c, neutrophils alone were sufficient to enhance tumor metastasis to lung.

Neutrophils promote tumor metastasis. (a and b) For the wound-healing assay, cultured 4T1 cells were first starved for 24 h. New medium was added and freshly isolated neutrophils were added. Wound closures were photographed (a) and statistically analyzed (b) at 24 h after wounding (scale bars: 700 μm). (c) 4T1 breast cancer cells (4 × 105) alone or with freshly isolated neutrophils (4 × 105) were co-injected into fat pad of BALB/c mice. Mice were euthanized after 4 weeks. Metastasis of tumor nodules on lung was displayed by counting. (d and e) Animal experiments were as described in Figure 1a. SB265610 was i.p injected (2 mg/kg) to mice every days. Metastasis of tumor nodules on lung was displayed by counting (d), HE staining (scale bars: 700 μm) and whole-lung images (stained by bouin’s solution) (e). (f) Animal survival analysis. BALB/c mice were injected with 4T1 cells alone or with different MSCs as described in Figure 1a. SB265610 was i.p injected (2 mg/kg) to mice every 2 days. Data are shown as means±s.e.m. Statistical significance was assessed by unpaired, two-tailed Student’s t-test.
Next, we co-injected tumor cells with different MSCs into mice. We then treated mice with the CXCR2 antagonist through intraperitoneal injection. SB265610 efficiently decreased neutrophil infiltration into tumor (Supplementary Figure S5b), but did not significantly change the numbers of neutrophil in the blood (Supplementary Figure S5c). On the other hand, quantitation of tumor nodules in the lung, and H&E staining of the lung tissues showed that SB265610 significantly decreased the pro-metastatic effect of TNFα-activated MSCs (Figures 5d and e), whereas no significant difference in primary tumor growth was observed (Supplementary Figure S5d). Furthermore, CXCR2 antagonist treatment also prolonged the survival time of tumor-bearing mice (Figure 5f). An Ly6G-neutralizing antibody was also used to specifically depleted neutrophils in mice. Indeed, we found that Ly6G antibody significantly decreased lung metastasis in the presence of TNFα-activated MSCs (Supplementary Figure S5e). Therefore, these data strongly suggest that neutrophils in the tumor have a critical role in tumor metastasis.
Neutrophils stimulate tumor cells to express high levels of pro-metastatic factors
There are a substantial number of studies that have suggested that neutrophils can promote tumor metastasis through the secretion of MMPs and selected growth factors, such as IL-6 and TGFβ.31, 32, 33 Meanwhile, the characteristics of tumor cells are also modified by neutrophils.34 Based on these observations, we co-cultured 4T1 cells with freshly isolated neutrophils in vitro and examined the expression of pro-metastatic factors by the tumor cells. After co-culture with neutrophils for 12 h, we found the mRNA levels of multiple genes were markedly elevated in 4T1 cells, including chemokine receptors (CXCR4, CXCR7), MMPs (MMP12, MMP13) and growth factors (IL-6, TGFβ) (Figure 6a). In addition, previous reports also suggested that S100A8/9 enhanced breast cancer survival and chemotherapy resistance.35 Indeed, we found a high level of S100A8/9 expressed by neutrophils, but not by MSCs or tumor cells (Figure 6b).

Neutrophils induce expression of pro-metastatic factors in tumor cells. (a) 4T1 cells were co-cultured with freshly isolated neutrophils for 12 h, after which time, the neutrophils were washed off of the plate and gene expression in 4T1 cells was detected by qPCR. (b) mRNA levels of S100A8/9 in different cells were detected by qPCR. (c and d) Overall survival (c) and distant metastasis-free survival (d) of breast cancer patients with high- or low- IL8 (Affymetrix ID: 202859_x_at) expression. Hazard ratios (HR) and P-values (log-rank P) were depicted for each survival analysis. Kaplan–Meier survival data were generated using the publicly online tool KM-plotter. (e) IL8 expression in TNFα-treated human MSCs (10 ng/ml, 12 h) was analyzed by real-time PCR. (f) Correlation analysis of IL8 and other pro-metastatic genes expression in breast cancer patient samples.
To investigate whether same mechanism exists in human breast cancer, we used an online database of potential breast cancer biomarkers (http://www.kmplot.com) to assess the prognostic implications of the expression of CXCR2 ligands in breast cancer patients. We examined all of the CXCR2 ligands and found that only IL8 levels showed correlations with breast cancer patient survival. Expression of high levels of IL8 was found to correlate with poor overall survival and distant metastasis-free survival (Figures 6c and d).
IL8, also known as CXCL8, is a well-studied chemotactic factor for neutrophils.36 The IL8 gene is observed in humans, but is absent in mouse and rat.37 After TNFα activation, human MSCs significantly upregulated IL8 levels (Figure 6e). We next investigated gene expression in a cohort of clinical breast cancer samples. Tumor specimens from 33 breast cancer patients were analyzed for gene expression. Based on the expression of IL8, the samples were divided into two groups. Interestingly, tumors with higher levels of IL8 expression tended to have much higher levels of pro-metastatic factors (Figure 6f). Therefore, our data suggested that neutrophil accumulation in human tumors may also result in promotion of tumor malignancy and metastasis.
Discussion
Tumor metastatic models have demonstrated that MSCs in primary tumors can influence tumor cell invasion and metastasis by providing contextual signals. Also, MSCs orchestrated the tumor immune microenvironment through their cross-talk with inflammatory cells.10, 13, 38 Here we demonstrate that TNFα-activated MSCs enhance breast cancer lung metastasis, and suggest that interactions between MSCs and inflammation endow the TME with more vigorous neutrophil infiltration, thereby promoting tumor metastasis.
Although the role of TNFα-activated MSCs in tumor progression has been shown in various types of cancer,13 there is currently no available evidence showing their effect on tumor metastasis. Our demonstration of increased metastatic nodules in the lungs of mice with 4T1 breast cancer cell-derived tumors, in the presence of injection of TNFα-activated MSCs, indicates that TNFα-activated MSCs promote breast cancer metastasis to the lung. Our previous study reported that TNFα-activated MSCs mimicked T-MSCs, with the comparable ability to promote tumor growth.13, 39 Similarly, we found that T-MSCs advanced tumor metastasis. Although a previous report showed that MSCs could promote the migration of mammary cancer cells in vitro,40 studies from our tumor model showed that TNFα-activated MSCs failed to directly enhance 4T1 cell migration, suggesting that MSCs enhanced tumor metastasis through their interaction with other types of cells in the TME. We further demonstrated that CD11b+Ly-6G+ cells were the major cell population involved in tumor metastasis enhanced by TNFα-activated MSCs. Previous studies have reported that CD11b+Gr1+ and other myeloid cell types can facilitate tumor growth and metastasis.41, 42, 43 CD11b+Gr1+ cells are a cell population comprised by neutrophils and monocytes.43 It has been reported that CCL2/CCR2 signaling mediated monocyte/macrophage accumulation facilitates breast cancer metastasis.21 However, we excluded the possibility that monocytes and macrophages were involved in the enhanced tumor metastasis by TNFα-activated MSCs. Only blockade of CXCR2 could abolish the advancement of tumor metastasis by TNFα-activated MSCs, indicating that CXCR2+ cells are the key cell population that mediates the pro-metastatic role of TNFα-activated MSCs. It has been demonstrated that CXCR2 ligands are critical in recruiting neutrophils in the process of inflammation.35, 44 Taken together, these data confirmed CXCR2+ neutrophil accumulation to be critical for the pro-metastatic microenvironment prepared by TNFα-activated MSCs.
Although increased neutrophil infiltration was observed in both the pro-metastatic tumor site and the pre-metastatic lung in breast cancer model,43, 45 CXCR2 ligands CXCL1, CXCL2 and CXCL5 exhibited high levels of expression in the primary tumor site, but not bone marrow or lung. This result indicates that tumor MSCs-induced neutrophil increase in the primary tumor site and blood has a critical role in regulating tumor metastasis. Consistently, removal of the primary tumor can lead to the reduction of neutrophils in peripheral blood.45 Our previous study pointed that MSCs could inhibit lymphocyte apoptosis through IL-6 secretion.46 Studies also found that IL-6 produced by human MSCs can suppress neutrophil apoptosis.47 The supporting effect of TNFα-activated MSCs on neutrophil survival will be of interest to study in further experiments.
The role of neutrophils in the steps of tumor metastasis was also studied. Some genes, such as S100A8 and S100A9, were increased in neutrophils that were related to tumor metastasis.35 We also observed high levels of S100A8 and S100A9 expression in neutrophils, but not stromal cells and tumor cells. However, the function of these neutrophil-derived factors remains elusive. Our data have demonstrated that tumor cells co-cultured with neutrophils express higher levels of pro-metastatic genes, including CXCR4, CXCR7, MMP12, MMP13, IL-6 and TGFβ. In turn, these tumor cells might shape the TME, even affecting the properties of tumor-infiltrating neutrophils. Interestingly, in comparison of blood-derived neutrophils and tumor-derived neutrophils, we found that tumor-derived neutrophils showed elevated expression of multiple growth factors (for example, IL-6, VEGF), compared with that of blood-derived neutrophils (data not shown). Moreover, those tumor-derived neutrophils resulted in more severe lung metastasis than blood-derived neutrophils (data not shown). A comparison of tumor-associated neutrophils, granulocytic myeloid-derived suppressor cells and normal neutrophils by transcriptomic analysis found that the TME influenced the mRNA program of neutrophils, with a prominent feature to promote tumor progression.48 The molecular mechanisms modifying neutrophils during tumor metastasis will be an important area for study. For instance, whether neutrophil-secreted factors affect tumor stem cell stemness through targeting oncogenes such as c-myc49 and p6350, 51, 52 is important to understand. Nevertheless, these results indicated that neutrophils infiltration of the TME could be the result of a series of events initiated by the inflammatory cytokine activated MSCs.
In conclusion, our findings provide a novel mechanism through which MSCs promote tumor metastasis. CXCR2+ neutrophil accumulation in pro-metastatic microenvironment is induced by the regulatory effects of TNFα on MSCs. These neutrophils can directly enhance tumor lung metastasis by enhancing tumor cell expression of pro-metastatic genes. Therefore, targeting MSC-induced immune cell chemotaxis may provide a potential method for clinical cancer therapy.
Materials and methods
Mice
BALB/c mice were purchased from the Shanghai Laboratory Animal Center of Chinese Academy of Sciences, Shanghai, China, and maintained under specific pathogen-free conditions. Mice were maintained in the vivarium of Shanghai Jiao Tong University School of Medicine. Animals were matched for age and gender in each experiment. The animal protocols for the experiments described in this paper were approved by the Institutional Animal Care and Use Committee of the Institute of Health Sciences, Shanghai Institutes for Biological Sciences of Chinese Academy of Sciences.
Cells
MSCs were generated from tibia and femur bone marrow or by isolation from lung and tumor digested with type II collagenase (Sigma-Aldrich, St Louis, MO, USA) using 6–10 week-old mice. Cells were cultured in DMEM supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin (all from Invitrogen, Carlsbad, CA, USA). Cells were used before 20th passage. The identity of the MSCs was confirmed by their capability to differentiate into adipocytes, osteoblasts and chondrocytes, and by their expression of specific cell surface markers.
Reagents
Recombinant mouse TNFα, ELISA kit for testing mouse CCL5, CXCL1 and CXCL5 were purchased from R&D Systems (Minneapolis, MN, USA). FACS antibodies were from eBiosciences (La Jolla, CA, USA). Bouin’s solution, RS504393 and SB431542 were from Sigma-Aldrich. Mouse neutrophil isolation kit was purchased from Miltenyi Biotec (Bergisch Gladbach, Germany). Ly6G antibody and isotype antibody were purchased from Biolegend (San Diego, CA, USA). PCR Array Kit for Mouse Chemokines & Receptors was purchased from SABiosciences (Valencia, CA, USA).
Animal studies
MSCs (1 × 105) were co-injected with 4T1 tumor cells (4 × 105) into BALB/c mice in the intramammary gland fat pad. At various time points, tumor sizes were measured and the immune cell frequency in tumors or peripheral blood was analyzed by flow cytometry. Four weeks after tumor cell administration, the resultant tumors were excised and weighed. Tumor nodes on lung were counted and showed by HE staining.
Real-Time PCR
Total RNA was isolated using RNAprep pure Cell/Bacteria Kit (Tiangen Biotech, Beijing, China), and reverse-transcribed into complementary DNA was performed using first complementary DNA Synthesization Kit with oligo(dT)15 (Tiangen Biotech). The levels of mRNA of genes of interest were measured by real-time PCR (7900 HT by Applied Biosystems, Foster City, CA, USA) using SYBR Green Master Mix (Roche Diagnostics, Indianapolis, IN, USA). Total amount of mRNA was normalized to endogenous β-actin mRNA. Sequences of PCR primer pairs were listed in Supplementary Table 1.
Flow cytometric analysis
Cells surface markers were stained according to the R&D flow cytometry protocol. In brief, cells were firstly harvested and washed with PBS. Cells were then resuspended in PBS containing 0.5% bovine serum albumin and were incubated on ice for 30 min with fluorochrome-conjugated antibodies. Then stained cells were washed and analyzed on a FACS Calibur flow cytometer (Becton Dickinson, SanJose, CA, USA). FCS Express software was used for data analysis.
Clinical samples
The tumor specimens from breast cancer patients in this study were obtained from the Tumor Center of the Institute of Health Sciences, Shanghai Institutes for Biological Sciences. All individuals provided informed consent. The study was approved by the institutional biomedical research ethics committee of the Shanghai Institutes for Biological Sciences (Chinese Academy of Sciences).
Statistical Analysis
For statistical analysis, mean values with s.e.m. were presented in most graphs that were derived from several repeats of biological experiments. Statistical significance was assessed by unpaired two-tailed Student’s t-test. The Kaplan–Meier method was used to estimate the survival curves for the patient. For animal survival analysis, log-rank (Mantel–Cox) test was used.
Abbreviations
- MSCs:
-
mesenchymal stromal cells
- T-MSCs:
-
tumor-derived MSCs
- TNFα:
-
tumor necrosis factor α
- TME:
-
tumor microenvironment
- TANs:
-
tumor-associated neutrophils
References
Ohnuki H, Jiang K, Wang D, Salvucci O, Kwak H, Sanchez-Martin D et al. Tumor-infiltrating myeloid cells activate Dll4/Notch/TGF-beta signaling to drive malignant progression. Cancer Res 2014; 74: 2038–2049.
Shaked Y, Henke E, Roodhart JM, Mancuso P, Langenberg MH, Colleoni M et al. Rapid chemotherapy-induced acute endothelial progenitor cell mobilization: implications for antiangiogenic drugs as chemosensitizing agents. Cancer Cell 2008; 14: 263–273.
Ho IA, Chan KY, Ng WH, Guo CM, Hui KM, Cheang P et al. Matrix metalloproteinase 1 is necessary for the migration of human bone marrow-derived mesenchymal stem cells toward human glioma. Stem Cells 2009; 27: 1366–1375.
Bianco P, Robey PG, Simmons PJ . Mesenchymal stem cells: revisiting history, concepts, and assays. Cell Stem Cell 2008; 2: 313–319.
Sasaki M, Abe R, Fujita Y, Ando S, Inokuma D, Shimizu H . Mesenchymal stem cells are recruited into wounded skin and contribute to wound repair by transdifferentiation into multiple skin cell type. J Immunol 2008; 180: 2581–2587.
Wu Y, Chen L, Scott PG, Tredget EE . Mesenchymal stem cells enhance wound healing through differentiation and angiogenesis. Stem Cells 2007; 25: 2648–2659.
Hanahan D, Weinberg RA . Hallmarks of cancer: the next generation. Cell 2011; 144: 646–674.
Dwyer RM, Potter-Beirne SM, Harrington KA, Lowery AJ, Hennessy E, Murphy JM et al. Monocyte chemotactic protein-1 secreted by primary breast tumors stimulates migration of mesenchymal stem cells. Clin Cancer Res 2007; 13: 5020–5027.
Rattigan Y, Hsu JM, Mishra PJ, Glod J, Banerjee D . Interleukin 6 mediated recruitment of mesenchymal stem cells to the hypoxic tumor milieu. Exp Cell Res 2010; 316: 3417–3424.
Karnoub AE, Dash AB, Vo AP, Sullivan A, Brooks MW, Bell GW et al. Mesenchymal stem cells within tumour stroma promote breast cancer metastasis. Nature 2007; 449: 557–563.
Huang Y, Yu P, Li W, Ren G, Roberts AI, Cao W et al. p53 regulates mesenchymal stem cell-mediated tumor suppression in a tumor microenvironment through immune modulation. Oncogene 2014; 33: 3830–3838.
Lourenco S, Teixeira V, Kalber T, Jose R, Floto R, Janes S . Macrophage migration inhibitory factor–CXCR4 is the dominant chemotactic axis in human mesenchymal stem cell recruitment to tumors. J Immunol 2015; 194: 3463–3474.
Ren G, Zhao X, Wang Y, Zhang X, Chen X, Xu C et al. CCR2-dependent recruitment of macrophages by tumor-educated mesenchymal stromal cells promotes tumor development and is mimicked by TNFalpha. Cell Stem Cell 2012; 11: 812–824.
Chaturvedi P, Gilkes DM, Takano N, Semenza GL . Hypoxia-inducible factor-dependent signaling between triple-negative breast cancer cells and mesenchymal stem cells promotes macrophage recruitment. Proc Natl Acad Sci 2014; 111: E2120–E2129.
Turley SJ, Cremasco V, Astarita JL . Immunological hallmarks of stromal cells in the tumour microenvironment. Nat Rev Immunol 2015; 15: 669–682.
Ren G, Zhang L, Zhao X, Xu G, Zhang Y, Roberts AI et al. Mesenchymal stem cell-mediated immunosuppression occurs via concerted action of chemokines and nitric oxide. Cell Stem Cell 2008; 2: 141–150.
Han X, Yang Q, Lin L, Xu C, Zheng C, Chen X et al. Interleukin-17 enhances immunosuppression by mesenchymal stem cells. Cell Death Differ 2014; 21: 1758–1768.
Li W, Ren G, Huang Y, Su J, Han Y, Li J et al. Mesenchymal stem cells: a double-edged sword in regulating immune responses. Cell Death Differ 2012; 19: 1505–1513.
Kitamura T, Qian B-Z, Pollard JW . Immune cell promotion of metastasis. Nat Rev Immunol 2015; 15: 73–86.
Yang P, Li Q-J, Feng Y, Zhang Y, Markowitz GJ, Ning S et al. TGF-β-miR-34a-CCL22 signaling-induced Treg cell recruitment promotes venous metastases of HBV-positive hepatocellular carcinoma. Cancer Cell 2012; 22: 291–303.
Qian B-Z, Li J, Zhang H, Kitamura T, Zhang J, Campion LR et al. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 2011; 475: 222–225.
Jochems C, Schlom J . Tumor-infiltrating immune cells and prognosis: the potential link between conventional cancer therapy and immunity. Exp Biol Med 2011; 236: 567–579.
Zhang Q-w, Liu L, Gong C-y, Shi H-s, Zeng Y-h, Wang X-z et al. Prognostic significance of tumor-associated macrophages in solid tumor: a meta-analysis of the literature. PLoS One 2012; 7: e50946.
Templeton AJ, McNamara MG, Šeruga B, Vera-Badillo FE, Aneja P, Ocaña A et al. Prognostic role of neutrophil-to-lymphocyte ratio in solid tumors: a systematic review and meta-analysis. J Natl Cancer Inst 2014; 106: dju124.
Gregory AD, Houghton AM . Tumor-associated neutrophils: new targets for cancer therapy. Cancer Res 2011; 71: 2411–2416.
Fridlender ZG, Albelda SM . Tumor-associated neutrophils: friend or foe? Carcinogenesis 2012; 33: 949–955.
Zhou SL, Dai Z, Zhou ZJ, Chen Q, Wang Z, Xiao YS et al. CXCL5 contributes to tumor metastasis and recurrence of intrahepatic cholangiocarcinoma by recruiting infiltrative intratumoral neutrophils. Carcinogenesis 2014; 35: 597–605.
Gijsbers K, Gouwy M, Struyf S, Wuyts A, Proost P, Opdenakker G et al. GCP-2/CXCL6 synergizes with other endothelial cell-derived chemokines in neutrophil mobilization and is associated with angiogenesis in gastrointestinal tumors. Exp Cell Res 2005; 303: 331–342.
Bald T, Quast T, Landsberg J, Rogava M, Glodde N, Lopez-Ramos D et al. Ultraviolet-radiation-induced inflammation promotes angiotropism and metastasis in melanoma. Nature 2014; 507: 109–113.
De Luca A, Lamura L, Gallo M, Maffia V, Normanno N . Mesenchymal stem cell-derived interleukin-6 and vascular endothelial growth factor promote breast cancer cell migration. J Cell Biochem 2012; 113: 3363–3370.
Ardi VC, Kupriyanova TA, Deryugina EI, Quigley JP . Human neutrophils uniquely release TIMP-free MMP-9 to provide a potent catalytic stimulator of angiogenesis. Proc Natl Acad Sci 2007; 104: 20262–20267.
Yang L, Huang J, Ren X, Gorska AE, Chytil A, Aakre M et al. Abrogation of TGF beta signaling in mammary carcinomas recruits Gr-1+CD11b+ myeloid cells that promote metastasis. Cancer Cell 2008; 13: 23–35.
Yan B, Wei J-J, Yuan Y, Sun R, Li D, Luo J et al. IL-6 cooperates with G-CSF to induce protumor function of neutrophils in bone marrow by enhancing STAT3 activation. J Immunol 2013; 190: 5882–5893.
Hu P, Shen M, Zhang P, Zheng C, Pang Z, Zhu L et al. Intratumoral neutrophil granulocytes contribute to epithelial-mesenchymal transition in lung adenocarcinoma cells. Tumour Biol 2015; 36: 7789–7796.
Acharyya S, Oskarsson T, Vanharanta S, Malladi S, Kim J, Morris PG et al. A CXCL1 paracrine network links cancer chemoresistance and metastasis. Cell 2012; 150: 165–178.
Kunkel S, Standiford T, Kasahara K, Strieter R . Interleukin-8 (IL-8): the major neutrophil chemotactic factor in the lung. Exp Lung Res 1991; 17: 17.
Modi W, Yoshimura T . Isolation of novel GRO genes and a phylogenetic analysis of the CXC chemokine subfamily in mammals. Mol Biol Evol 1999; 16: 180–193.
El-Haibi CP, Karnoub AE . Mesenchymal stem cells in the pathogenesis and therapy of breast cancer. J Mammary Gland Biol Neoplasia 2010; 15: 399–409.
Ren G, Liu Y, Zhao X, Zhang J, Zheng B, Yuan ZR et al. Tumor resident mesenchymal stromal cells endow naive stromal cells with tumor-promoting properties. Oncogene 2013; 33: 4016–4020.
Halpern J, Kilbarger A, Lynch C . Mesenchymal stem cells promote mammary cancer cell migration in vitro via the CXCR2 receptor. Cancer Lett 2011; 308: 91–99.
Yang L, DeBusk LM, Fukuda K, Fingleton B, Green-Jarvis B, Shyr Y et al. Expansion of myeloid immune suppressor Gr+ CD11b+ cells in tumor-bearing host directly promotes tumor angiogenesis. Cancer Cell 2004; 6: 409–421.
Shojaei F, Wu X, Malik AK, Zhong C, Baldwin ME, Schanz S et al. Tumor refractoriness to anti-VEGF treatment is mediated by CD11b+ Gr1+ myeloid cells. Nat Biotechnol 2007; 25: 911–920.
Kowanetz M, Wu X, Lee J, Tan M, Hagenbeek T, Qu X et al. Granulocyte-colony stimulating factor promotes lung metastasis through mobilization of Ly6G+ Ly6C+ granulocytes. Proc Natl Acad Sci 2010; 107: 21248–21255.
Katoh H, Wang D, Daikoku T, Sun H, Dey SK, Dubois RN . CXCR2-expressing myeloid-derived suppressor cells are essential to promote colitis-associated tumorigenesis. Cancer Cell 2013; 24: 631–644.
Coffelt SB, Kersten K, Doornebal CW, Weiden J, Vrijland K, Hau C-S et al. IL-17-producing [ggr][dgr] T cells and neutrophils conspire to promote breast cancer metastasis. Nature 2015; 522: 345–348.
Xu G, Zhang Y, Zhang L, Ren G, Shi Y . The role of IL-6 in inhibition of lymphocyte apoptosis by mesenchymal stem cells. Biochem Biophys Res Commun 2007; 361: 745–750.
Raffaghello L, Bianchi G, Bertolotto M, Montecucco F, Busca A, Dallegri F et al. Human mesenchymal stem cells inhibit neutrophil apoptosis: a model for neutrophil preservation in the bone marrow niche. Stem Cells 2008; 26: 151–162.
Fridlender ZG, Sun J, Mishalian I, Singhal S, Cheng G, Kapoor V et al. Transcriptomic analysis comparing tumor-associated neutrophils with granulocytic myeloid-derived suppressor cells and normal neutrophils. PloS One 2012; 7: e31524.
Wang J, Wang H, Li Z, Wu Q, Lathia J, Mclendon R et al. c-Myc is required for maintenance of glioma cancer stem cells. PloS One 2008; 3: e3769.
Memmi E, Sanarico A, Giacobbe A, Peschiaroli A, Frezza V, Cicalese A et al. p63 sustains self-renewal of mammary cancer stem cells through regulation of Sonic Hedgehog signaling. Proc Natl Acad Sci USA 2015; 112: 3499–3504.
Giacobbe A, Compagnone M, Bongiornoborbone L, Antonov A, Markert E, Zhou J et al. p63 controls cell migration and invasion by transcriptional regulation of MTSS1. Oncogene 2015; 35: 1602–1608.
Melino G, Memmi E, Pelicci P, Bernassola F . Maintaining epithelial stemness with p63. Sci Signal 2015; 8: re9.
Acknowledgements
This study was supported by grants from the Scientific Innovation Project of the Chinese Academy of Sciences (XDA 01040100), the Ministry of Science and Technology of China (2015CB964400), Major Program of Development Fund for Shanghai Zhangjiang National Innovation Demonstration Zone (ZJ2014-ZD-002), the Programs of National Natural Science of China (81330046, 81273316, 81571612, 81530043), the External Cooperation Program of BIC, Chinese Academy of Sciences (GJHZ201307), Shanghai Rising-Star Program (14QA1404200) and Youth Innovation Promotion Association, Chinese Academy of Sciences.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on the Oncogene website
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Yu, P., Huang, Y., Han, Y. et al. TNFα-activated mesenchymal stromal cells promote breast cancer metastasis by recruiting CXCR2+ neutrophils. Oncogene 36, 482–490 (2017). https://doi.org/10.1038/onc.2016.217
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2016.217
This article is cited by
-
Neutrophils in the premetastatic niche: key functions and therapeutic directions
Molecular Cancer (2024)
-
Prognostic marker CXCL5 in glioblastoma polyformis and its mechanism of immune invasion
BMC Cancer (2024)
-
Understanding the role of TNFR2 signaling in the tumor microenvironment of breast cancer
Journal of Experimental & Clinical Cancer Research (2024)
-
Neutrophils in cancer: dual roles through intercellular interactions
Oncogene (2024)
-
Pre-metastatic niche: formation, characteristics and therapeutic implication
Signal Transduction and Targeted Therapy (2024)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}