
Abstract
Of the many dendritic cell (DC) subsets, DCs expressing the monomorphic coreceptor CD8 α-chain (CD8α) are localized permanently in lymphoid organs, whereas 'tissue-derived DCs' remain in nonlymphoid tissues until they 'capture' antigen and then move to local lymph nodes. Here we show that after lung infection, both naive and memory CD8+ 'killer' T cells responded to influenza virus antigens presented by lymph node–resident CD8α+ DCs, but only naive cells responded to antigens presented by lung-derived DCs. This difference provides a mechanism for priming naive T cell responses in conditions in which robust memory predominates. Our findings have implications for immunity to pathogens that can mutate their T cell epitopes, such as influenza virus and human immunodeficiency virus, and challenge the long-held view that memory T cells have less-stringent requirements for activation than naive T cells have.
Similar content being viewed by others
Main
The generation of infection-fighting killer CD8-expressing (CD8+) T cells, responsible for destroying virus-infected cells, requires antigen presentation by dendritic cells (DCs)1,2,3. The classic model of DC function suggests that immature DCs are located in almost all tissues of the body, where they screen the environment for pathogens4. In these tissues, immature DCs efficiently capture antigens but poorly express those molecules necessary for T cell priming. After pathogen encounter, however, DCs undergo a maturation program that involves their migration to the lymph node and the upregulation of molecules essential for T cell priming. Although this model has been explanatorily helpful, it does not encompass data suggesting that the DC network is increasingly more complex4,5,6.
DCs can be divided into distinct subsets: the first key division is that between plasmacytoid DCs and conventional DCs7. Plasmacytoid DCs differ from conventional DCs in several ways but most notably in their dominant capacity to synthesize interferon-α in response to exogenous stimuli such as viral particles8. Conventional DCs can be further divided into many subsets6,9,10, of which only a relatively shallow understanding of subset-specific functions now exists. In the steady state, there seem to be at least five different conventional DC subtypes in lymph nodes10, three of which are also found in the spleen9. The three subsets found in both the spleen and lymph nodes are derived from precursors that enter from the blood11 and are essentially lymphoid tissue–resident DCs. In contrast, two additional DC subtypes found in lymph nodes but not the spleen access the former through migration from peripheral tissues12; these DCs consist of Langerhans cells and dermal DCs of the skin, which are essentially the interstitial counterparts of dermal DCs found in other tissues. In addition to these various conventional DC subsets found in the steady state, monocyte-derived DCs contribute to immunity in inflammatory conditions13.
Studies examining the function of DC subsets in the initiation of immunity to viral infection have shown that lymphoid tissue–resident CD8α+ DCs are heavily involved in priming CD8+ killer T cell responses14,15,16,17,18, regardless of whether the infection route is the blood, the skin or the lungs17,18. In contrast to the classic DC model, however, CD8α+ DCs seem to capture their viral antigen from other DCs that migrate from the site of infection (lung or skin) into the draining lymph nodes18,19. During influenza virus infection of the lungs, two subsets of DCs present viral antigens to naive CD8+ T cells18: lung-derived migratory DCs (CD205+CD11b−CD8α−), which transport antigen to the lymph node; and lymph node–resident CD8α+ DCs (CD205+CD11b−CD8α+), which 'receive' antigen from the lung-derived CD8α− DCs. Such transfer of antigen from migratory lung-derived CD8α− DCs to lymph node–resident CD8α+ DCs seems to provide a means for amplifying the number of DCs in the lymph node that present antigen, thereby improving the likelihood of antigen-specific DC–T cell interactions20.
Although the interaction between lung-derived migratory CD8α− DCs and lymph node–resident CD8α+ DCs in response to lung infection with influenza virus already seems to be more complex than previously envisaged, present knowledge of DC subsets in the lung-draining lymph node suggests that at least three more DC subsets in this lymph node could potentially participate in immunity. These are plasmacytoid DCs and two additional lymph node–resident conventional DC subsets: CD11b+CD205−CD4− DCs and CD11b+CD205−CD4+ DCs. Because memory CD8+ T cells have been reported to have less-stringent requirements for activation than naive T cells have21,22, we examined here whether these additional DC subsets might be capable of initiating secondary responses by memory T cells. We found that, unexpectedly, memory T cells not only responded poorly to the additional DC subsets but also were poorly responsive to migratory lung-derived CD8α− DCs, despite the ability of the latter to stimulate naive T cells. Our data provide insight into the function of migratory CD8α− DCs in priming naive T cells and indicate involvement of lymph node–resident CD8α+ DCs in the generation of secondary responses.
Results
Poor memory responses to lung-derived CD8α− DCs
To determine whether memory T cells respond to DC subsets other than lung-derived CD8α− DCs and lymph node–resident CD8α+ DCs, which have been shown to activate naive T cells during lung infection with influenza virus, we first generated memory T cells in vitro by stimulating naive T cell receptor–transgenic CD8+ T cells with antigen and then culturing them for at least 14 d with interleukin 15 (IL-15)23 (Supplementary Fig. 1 online). We then used these cells as responders to stimulation by various DC subtypes isolated ex vivo from the lung-draining lymph nodes of virus-infected mice. In these experiments, we used recombinant WSN influenza virus expressing a major histocompatibility complex class I–restricted epitope of herpes simplex virus glycoprotein B ((gB)24; called 'WSN-gB' here) as the source of antigen and naive or memory T cells from gB-specific T cell receptor–transgenic mice (gBT-I mice) as responding T cell populations. At 3 d after intranasal infection of C57BL/6 (B6) mice, corresponding to the peak of antigen presentation18, we prepared CD11c+ DCs from mediastinal lymph nodes by depleting the lymph nodes of various cells, including plasmacytoid DCs, and separated the cells into subsets by flow cytometry gated on CD11b and CD8α expression. Lung-derived DCs are CD11b−CD8α−, whereas lymph node–resident DCs, responsible for presenting viral antigens to naive T cells, are CD11b−CD8α+ (ref. 18). The remaining CD11b+ DCs are poorly defined, but most probably represent other types of lymph node–resident DC.
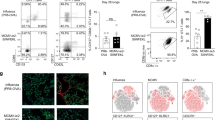
We found that, unexpectedly, memory CD8+ gBT-I T cells were less broadly responsive to stimulation by the various DCs than were naive gBT-I cells (Fig. 1a). As shown by the percent and number of proliferating T cells, the memory T cells did not proliferate in response to antigen presented by lung-derived CD8α− DCs, although they remained reactive to antigen presented by lymph node–resident CD8α+ DCs. We obtained the same results when we used either memory gBT-I cells primed in vivo by viral infection (Supplementary Fig. 2 online) or 'authentic' endogenous memory T cells generated in vivo by virus infection (Fig. 1b). In the latter experiment, we separated DCs on the basis of the expression of CD45RA and CD8: lung-derived CD8α− DCs were in the double-negative group of DCs and plasmacytoid DCs were represented by CD45RA+ DCs. These data support the conclusion that memory CD8+ T cells are poorly stimulated by lung-derived CD8α− DCs regardless of the method used to generate memory T cells.

(a) Proliferation of CFSE-labeled naive or memory CD8+ gBT-I T cells after stimulation for 60 h with CD8α+CD11b− DCs (lymph node–resident CD8α+ DCs), CD8α−CD11b− DCs (lung-derived CD8α− DCs) or CD8α−CD11b+ DCs isolated from the mediastinal lymph nodes of mice 3 d after intranasal infection with WSN-gB. Numbers in top left corners indicate percent (and number, in parentheses) of proliferated cells. Data are representative of four experiments. (b) Proliferation of CFSE-labeled endogenous influenza-specific memory CD8+ T cells (CD62LhiCD44hi T cells from spleens of B6 mice primed with WSN-gB (top), or PR8 followed by HKx31 (bottom)) after stimulation for 60 h by CD8α+ DCs, CD8α−CD45RA− DCs (double-negative (DN) DCs) or CD45RA+ DCs (plasmacytoid DCs (pDCs)) isolated from the mediastinal lymph nodes of mice 3 d after intranasal infection with WSN-gB (top) or HKx31 (bottom) influenza virus. CD8+ T cells were stained with tetramers specific for H-2Kb-restricted gB(498–505) (KbgB(498-505); top) or the H-2Db-restricted nucleoprotein peptide of amino acids 366–374 (DbNP(366–374); bottom). Numbers in outlined areas indicate percent proliferated tetramer-positive cells. Data are representative of one of two experiments.
Examination of the time course of ex vivo antigen presentation by DCs to naive T cells after influenza virus infection showed that whereas lymph node–resident CD8α+ DCs presented viral antigens for the first 7 d, lung-derived CD8α− DCs (in the double-negative DC group; Fig. 1b) presented antigens for at least 9 d, as shown by the percent and number of proliferating T cells (Fig. 2a). If, as suggested by the data presented above, memory T cells can respond only to lymph node–resident CD8α+ DCs, then naive T cells but not memory T cells should respond in vivo at time points later than day 7 when only lung-derived CD8α− DCs will be presenting viral antigens. To test that hypothesis, we infected mice intranasally with WSN-gB and then, after 10 d, injected the mice with carboxyfluorescein diacetate succinimidyl diester (CFSE)–labeled naive or in vitro–generated memory CD8+ gBT-I T cells and examined the T cell proliferation that resulted in vivo 3 d later (Fig. 2b,c). As a positive control, we also injected CFSE-labeled T cells into other mice 3 d after infection with WSN-gB, a time when lymph node–resident CD8α+ DCs should be able to stimulate both naive and memory CD8+ T cells. Consistent with our hypothesis, naive and memory CD8+ T cells proliferated when injected into mice on day 3 of WSN-gB infection (Fig. 2c, bottom), but only the naive T cells proliferated at day 10 of infection (Fig. 2c, top). These data confirmed our ex vivo findings, showing that in vivo, the lung-derived CD8α− DCs failed to stimulate memory but not naive CD8+ T cells. The ability of memory T cells to respond on day 3 of infection (Fig. 2c, bottom) and the detection of these cells in the lymph node after transfer on day 10 of infection (Fig. 2c, middle) confirmed that the memory T cells had homed to lymph nodes.

(a) Proliferation of CFSE-labeled naive gBT-I T cells incubated for 60 h with CD45RA+ DCs (pDCs) or CD45RA− DCs that were either CD8α+ (lymph node–resident CD8α+ DC) or CD8α− (DN DCs; containing lung-derived CD8α− DCs) isolated from the mediastinal lymph nodes at various times after infection. Numbers in top left corners indicate percent (and number, in parentheses) of proliferated cells. Data are from one representative of two experiments with similar results (20 donor mice per time point; analyses at 3, 5, 7 and 9 d after infection were done in the same experiment). (b) Experimental protocol for c (above) and the extent of antigen presentation to T cells by purified DC subsets (below), as determined by direct ex vivo analysis in a. (c) Proliferation of CFSE-labeled naive or in vitro–generated memory CD8+ gBT-I T cells transferred into uninfected mice (middle), or mice infected 10 d (top) or 3 d (bottom) earlier with WSN-gB; proliferation of gBT-I T cells from the mediastinal lymph node was analyzed 3 d after transfer. Data are representative of five experiments with similar results.
Delineating the basis for poor memory responses
A trivial explanation for our observations is that memory CD8+ T cells kill lung-derived CD8α− DCs or suppress their function, thereby preventing them from inducing proliferation. If that were true, then culture of naive and memory T cells together with lung-derived CD8α− DCs should also abrogate proliferation of the naive population. We found, however, that naive T cells proliferated in those conditions (Fig. 3a), and, in control cultures, both populations responded as expected to lymph node–resident CD8α+ DCs (Fig. 3b). Conversely, to determine whether the poor response of memory CD8+ T cells to lung-derived CD8α− DCs could be explained by suppressive factors provided by the CD8α− DCs themselves, we compared the response of memory T cells to lymph node–resident CD8α+ DCs in the presence or absence of lung-derived CD8α− DCs (Fig. 3c). Lung-derived CD8α− DCs did not impair responses to lymph node–resident CD8α+ DCs, suggesting that the CD8α− DCs were not acting through a dominant suppressive mechanism.

(a,b) Proliferation of mixed cultures of naive and memory T cells cultured with lung-derived CD8α− DCs (a) or lymph node–resident CD8α+ DCs (b) from mice infected 3 d earlier with WSN-gB. Data are representative of two experiments. (c) Proliferation of CFSE-labeled naive or memory gBT-I cells cultured with lymph node–resident CD8α+ DCs, lung-derived CD8α− DCs or a mixture of equal numbers of each DC subtype. Data are representative of two independent experiments. (d,e) Proliferation of CFSE-labeled naive or memory gBT-I cells stimulated with peptide-coated lymph node–resident CD8α+ DCs or lung-derived CD8α− DCs from mediastinal lymph nodes of naive mice (d) or of peptide-coated lymph node–resident CD8α+ DCs and skin-derived CD8α− DCs (Langerhans cells and dermal DCs) from skin-draining lymph nodes of naive mice (e). DCs (5 × 103) were incubated for 60 min with 'titrating' concentrations of gB(498–505), were washed three times and then were cultured for 60 h with 5 × 104 CFSE-labeled gBT-I cells. Data are one representative of two experiments.
To assess more quantitatively the difference in the stimulatory capacity of lung-derived DCs for naive and memory T cells, we isolated both lymph node–resident CD8α+ DCs and lung-derived CD8α− DCs from naive mice, incubated them with various concentrations of gB peptide in vitro and then examined their ability to stimulate naive and memory CD8+ gBT-I T cells (Fig. 3d). Whereas naive T cells respond equivalently to lymph node–resident CD8α+ and lung-derived CD8α− DCs, memory T cells required ten-fold more peptide on lung-derived CD8α− DCs than on lymph node–resident CD8α+ DCs for an equivalent response. We obtained similar results when we used DCs from virus-infected mice (Supplementary Fig. 3 online). Thus, although lung-derived CD8α− DCs did not stimulate memory T cells to respond to influenza virus antigen during infection, this lack of stimulation did not represent a complete failure of lung-derived CD8α− DCs to activate memory T cells but instead represented a reduced capacity to stimulate them. Realistically, however, this reduction might mean that most natural stimuli are ineffective at stimulating memory T cells when presented on lung-derived CD8α− DCs. These findings extended beyond lung-derived CD8α− DCs; they were reproduced by comparison of peptide presentation by skin-derived DCs (Fig. 3e). Again, whereas lymph node–resident CD8α+ DC and skin-derived CD8α− DCs (consisting of a mixture of dermal DCs and Langerhans cells) stimulated naive T cells equivalently, skin-derived CD8α− DCs were less efficient than lymph node–resident CD8α+ DCs at stimulating memory T cells.
Naive responses are induced in the presence of memory
We considered that such a differential reaction of naive and memory T cells to tissue-derived DCs might exist to enable the response of new T cell specificities (from the naive repertoire) in conditions in which large numbers of competing memory T cells are already present. Such a response might be advantageous if pre-existing memory populations derived, for example, from cross-reactive infections were not particularly protective25,26. To investigate whether naive responses could still be primed when large numbers of competing memory cells were present, we assessed the in vivo population expansion of small numbers (5 × 104) of naive or memory gBT-I T cells (the 'responding population') in response to WSN-gB infection in the presence of 'titrated' numbers of naive or memory gBT-I cells (the 'competitor population'; Fig. 4a–d). We identified the responding population by its expression of a distinct Ly5 allotypic marker and assessed the number of cells generated in response to infection on day 10 (lung infection) or day 8 (intravenous infection).

(a–d) Flow cytometry of responding Ly5.1+ gBT-I cells in the spleens of B6 mice given adoptive transfer of 5 × 104 naive Ly5.1+ gBT-I cells (responders) and 'titrating' numbers of naive Ly5.2+ gBT-I cells (competitors; a), 5 × 104 memory Ly5.1+ gBT-I cells (responders) and 'titrating' numbers of naive Ly5.2+ gBT-I cells (competitors; b), or 5 × 104 naive Ly5.1+ gBT-I cells (responders) and titrating numbers of memory Ly5.2+ gBT-I cells (competitors; c,d), and then infected with WSN-gB either intranasally for 10 d (a–c) or intravenously for 8 d (d). Each line and symbol represents an individual experiment with at least two mice analyzed per data point. Data are from two to three experiments per condition. (e,f) Flow cytometry of antigen-specific CD8+ T cells from untreated mice (None) or mice given adoptive transfer of 2.2 × 106 CD44hiCD62hi memory CD8+ T cells (Memory cells) purified from mice that had been infected with influenza HKx31 at least 12 weeks earlier. At 24 h after T cell adoptive transfer, mice were infected with HKx31 intranasally for 10 d (e) or intravenously for 8 d (f), and then spleens were analyzed for the number of endogenous (e,f, left) or transferred memory CD8+ T cells specific for the H-2Db-restricted nucleoprotein peptide of amino acids 366–374 (DbNP(366)) or the H-2Db-restricted acid polymerase peptide of amino acids 224–233 (DbPA(224); e, right; f, middle). f, right, values for naive uninfected mice. Data are pooled from four (e) or two (f) experiments (each circle represents an individual mouse).
First, as a control, we showed that when in excess, naive T cells could outcompete other naive and memory T cells (Fig. 4a,b). Both outcomes were expected, because naive and memory T cells should be able to recognize antigen only on DCs that can also present to naive T cells. Notably, however, increasing numbers of memory T cells were very poor competitors against a naive responding population (Fig. 4c), although their presence was evident (Supplementary Fig. 4 online).
To support the idea that tissue-derived CD8α− DCs presented viral antigens to naive but not memory T cells (Fig. 4c), we examined competition of the same combination of T cells in circumstances in which tissue-derived CD8α− DCs were not involved. Intravenous viral infection results in presentation in the spleen by the lymphoid tissue–resident CD8α+ DCs alone17, which for lung infection was found to be capable of stimulating both memory and naive T cells (Fig. 1a). When we infected mice intravenously with WSN-gB (Fig. 4d), increasing numbers of memory T cells prevented naive responses, consistent with their ability to compete with naive T cells when the only DCs that are presenting viral antigens can stimulate both types of T cell.
To verify these findings with authentic (rather than transgenic) T cells, we isolated CD44hiCD62Lhi central memory T cells from B6.Ly5.1 mice at least 12 weeks after infection with HKx31 influenza virus and used these cells as competitors by adoptive transfer into B6 mice. We subsequently infected the B6 mice intranasally (Fig. 4e) or intravenously (Fig. 4f) with influenza virus and then examined the response by endogenous and transferred cells specific for viral nucleoprotein or viral acid polymerase. Consistent with our studies with transgenic T cells, memory CD8+ T cells prevented the response of naive endogenous T cells to influenza virus after intravenous infection but not after lung infection. These data collectively indicate that tissue-derived CD8α− DCs provide a 'preferential' avenue for naive T cell stimulation when competing memory cells are present. This explains why naive T cell responses have been detected despite the presence of preformed memory for lung infection with influenza virus27.
CD70 is used by lymph node–resident CD8α+ DCs
The data presented above also lead to other conclusions. We have shown that naive CD8+ T cells were more sensitive than memory CD8+ T cells in response to stimulation with tissue-derived CD8α− DCs and were equivalent to memory T cells in their response to lymph node–resident CD8α+ DCs (Figs. 1a, 2c and 3d,e). Those findings challenge the long-held model that memory T cells have fewer costimulatory requirements than naive T cells have, at least when tissue-derived DCs are used as antigen-presenting cells. How this difference is achieved at the molecular level is unclear, although we have excluded the possibility of obvious variations in the expression of various costimulatory molecules, including B7-H1, B7-H2, B7-DC, B7-RP, B7-1, B7-2 and BTLA-4 (Supplementary Fig. 5 online). On the basis of the reported diversity in DC subsets in their use of CD70 and IL-12 (ref. 28), we examined the function of these two molecules. The proliferative responses induced by lymph node–resident CD8α+ DCs were dependent on CD70 for both naive and memory T cells, whereas the stimulation of naive T cells by lung-derived CD8α− DCs was independent of either CD70 or IL-12 (Supplementary Fig. 6 online). This observation indicates that lung-derived CD8α− DCs use an alternative, as-yet-undefined costimulatory molecule to stimulate naive T cells efficiently, but this molecule stimulates memory T cells inefficiently.
Discussion
DCs are essential for the initiation of T cell responses to many infections1,29,30, emphasizing their central function in immunity. Despite the growing number of DC subsets identified in the spleen and lymph nodes6,9,10, extensive analysis has shown that only a few subsets are capable of priming virus-specific CD8+ T cells, at least for naive responses14,15,16,17,18,31,32. These subsets include CD8α+ DCs14,15,16,17,18,32, which seem to be lymphoid tissue resident15,18, and sometimes migratory CD8α− DCs18,31,32, particularly for lung infection with influenza virus18. Because DCs have also been shown to be essential for secondary T cell responses2,3 and memory T cells are reported to have less-stringent requirements for activation21,22, we postulated that additional DC subsets might be involved in the initiation of secondary T cell responses. Unexpectedly, however, memory CD8+ T cells showed a more limited capacity to respond to DC subsets after influenza virus infection of the lungs. Although naive T cells responded to both CD8α+ DCs and CD8α−CD11b−CD205+ migratory DCs shortly after infection, memory CD8+ T cell responses were restricted to CD8α+ DCs. The failure of migratory CD8α− DCs to activate memory CD8+ T cells was not absolute, as peptide coating enabled these DCs to stimulate memory T cells, although approximately tenfold more peptide was required for stimulation with CD8α− DCs than for stimulation with CD8α+ DCs. The poor capacity of CD8α− DCs to stimulate memory T cells extended to other migratory DCs, in particular those of the skin, suggesting that this observation applies more generally to migrating DCs.
Our findings indicate that memory T cells are highly dependent on antigen presentation by lymph node–resident CD8α+ DCs, as their capacity to respond to tissue-derived CD8α− DCs is limited. This difference in responsiveness may be important when an earlier viral infection generates weakly cross-reactive and nonprotective memory T cells. The mechanism that we have described here provides a means for circumventing restriction to a dominant but ineffective memory T cell specificity, as naive T cells with new specificities potentially capable of fighting infection will have an opportunity to be stimulated. The development of ineffective, cross-reactive memory T cells is probably rare for two different species of pathogen but more common for viruses such as human immunodeficiency virus33, lymphocytic choriomeningitis virus34, hepatitis C virus35 and even influenza virus36, which can mutate killer T cell epitopes.
This protective mechanism, which provides naive T cells with an avenue to respond in the face of dominant preformed memory, may also provide protection against dominant primary specificities that are nonprotective. If such dominant specificities are driven into memory as infection persists (and persistence would be a potential outcome when cytotoxic T lymphocytes are not protective), then subdominant naive T cells may be given a late opportunity to respond and to provide protection. This process might be particularly important for responses mediated by cross-presentation if, on occasion, a dominant cross-presented epitope is not well presented by the direct presentation pathway of infected cells.
Our studies here have emphasized differences in the ways that memory and naive CD8+ T cells interact with DC subsets, providing evidence that naive T cells may have fewer requirements than memory T cells have for activation, at least in the conditions that we have describe here. Our findings not only justify further scrutiny of the precise functions of individual DC subsets but also provide insight into new strategies for vaccine development. In prime-boost strategies, for example, targeting booster antigen to lymph node–resident CD8α+ DCs should be beneficial.
Methods
Mice.
B6 mice, B6.SJL-PtprcaPep3b/BoyJ (B6.Ly5.1) mice and gBT-I mice (on a B6 background)37 were from The Walter and Eliza Hall Institute of Medical Research animal facility and were maintained in specific pathogen–free conditions. Experiments began when mice were between 5 and 10 weeks of age and were done in accordance with guidelines of the Melbourne Directorate Animal Ethics Committee.
Virus infection.
Mice were anesthetized with methoxyfluorane and then were infected with a nonlethal challenge of recombinant influenza WSN-gB, which contains the gB(498–505) H-2Kb–restricted epitope of herpes simplex virus inserted into the neuraminidase stalk24. For intranasal infection, mice received 1 × 102.6 plaque-forming units (PFU) of WSN-gB or 1 × 104.5 PFU of HKx31 diluted in 25 μl PBS; for intravenous infection, mice received 1 × 102.95 PFU of WSN-gB diluted in 200 μl PBS or 1 × 105.3 PFU of HKx31 in 200 μl PBS; for intraperitoneal infection, mice received 1 × 107 PFU PR8 influenza virus in 500 μl PBS.
DC isolation and culture.
DC purification from the spleen or lymph nodes, flow cytometry and culture of DCs in vitro were done as described15,18,38,39. The number of DCs used varied between 6.25 × 103 and 12.5 × 103 cells per well for all analyses, except for peptide-coating experiments (5 × 103 DCs per well) and responses by endogenous memory T cells (5 × 104 DCs per well).
Preparation of CFSE-labeled CD8+ T cells.
Naive CD8+ gBT-I (H-2Kb-restricted anti-gB(498–505)) transgenic T cells were purified from pooled lymph nodes (inguinal, axillary, brachial, superficial cervical and mesenteric) by depletion of non-CD8+ T cells as described18. The T cell populations were routinely 85–95% CD8+Vα2+ as determined by flow cytometry. Naive and memory CD8+ T cells were labeled with CFSE39 or were used unlabeled. Proliferation was quantified after 60 h of culture. The gBT-I cells were labeled with CD8-specific monoclonal antibody and were resuspended in 100 μl balanced-salt solution plus 3% (vol/vol) FCS containing 2 × 104 'blank' calibration particles (BD Biosciences PharMingen). Samples were analyzed by flow cytometry on an LSR (Becton Dickinson), and the total number of live dividing lymphocytes (propidium iodide negative, CFSElo) was calculated from the number of dividing cells per 5 × 103 beads.
Generation of memory CD8+ T cell populations.
For the generation of in vitro–primed memory CD8+ T cells, an established model of the in vitro differentiation of central memory T cells was used3,23,40,41,42,43. Naive gBT-I transgenic spleen cells were coated for 1 h at 37 °C with 1 μM gB peptide. Cells were then washed twice in HEPES-buffered Earles medium containing 2.5% (vol/vol) FCS before being cultured at a density of 1.7 × 105 cells per ml in complete medium (mouse tonicity RPMI 1640 medium: RPMI 1640 medium containing 10% (vol/vol) FCS, 50 μM β-mercaptoethanol, 2 mM L-glutamine, 100 U/ml of penicillin and 100 μg/ml of streptomycin ('complete medium')). After 2 d, cells were washed and were supplemented with recombinant human IL-15 (20 ng/ml; R&D Systems). Complete medium containing human IL-15 was replaced every 3–4 d, and cells were used between 14 and 20 d after initiation of the culture.
For the generation of memory in vivo–primed CD8+ gBT-I T cells, 5 × 104 naive Ly5.1+ gBT-I T cells were adoptively transferred into Ly5.2+ congenic B6 mice. After 24 h, mice were infected with WSN-gB influenza virus. After 90 d, spleens from these mice were depleted of non-CD8+ cells and were stained with antibody to CD8 and antibody to Ly5.1. Memory Ly5.1+ CD8+ gBT-I T cells were then purified by flow cytometry. For the generation of authentic in vivo–primed influenza-specific memory T cells, mice either were primed first with PR8 intraperitoneally and then 8 weeks later with HKx31 intranasally or were primed with HKx31 alone or WSN-gB intranasally and left for at least 12 weeks.
Note: Supplementary information is available on the Nature Immunology website.
References
Jung, S. et al. In vivo depletion of CD11c+ dendritic cells abrogates priming of CD8+ T cells by exogenous cell-associated antigens. Immunity 17, 211–220 (2002).
Zammit, D.J., Cauley, L.S., Pham, Q.M. & Lefrancois, L. Dendritic cells maximize the memory CD8 T cell response to infection. Immunity 22, 561–570 (2005).
Belz, G.T. et al. Bone marrow-derived cells expand memory CD8+ T cells in response to viral infections of the lung and skin. Eur. J. Immunol. 36, 327–335 (2006).
Villadangos, J.A. & Heath, W.R. Life cycle, migration and antigen presenting functions of spleen and lymph node dendritic cells: limitations of the Langerhans cells paradigm. Semin. Immunol. 17, 262–272 (2005).
Heath, W.R. et al. Cross-presentation, dendritic cell subsets, and the generation of immunity to cellular antigens. Immunol. Rev. 199, 9–26 (2004).
Shortman, K. & Liu, Y. Mouse and human dendritic cell subtypes. Nat. Rev. Immunol. 2, 151–161 (2002).
Grouard, G. et al. The enigmatic plasmacytoid T cells develop into dendritic cells with interleukin (IL)-3 and CD40-ligand. J. Exp. Med. 185, 1101–1111 (1997).
Cella, M. et al. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat. Med. 5, 919–923 (1999).
Vremec, D., Pooley, J., Hochrein, H., Wu, L. & Shortman, K. CD4 and CD8 expression by dendritic cell subtypes in mouse thymus and spleen. J. Immunol. 164, 2978–2986 (2000).
Henri, S. et al. Hierarchy of susceptibility of dendritic cell subsets to infection by Leishmania major: inverse relationship to interleukin-12 production. Infect. Immun. 70, 3874–3880 (2002).
Shortman, K. & Naik, S.H. Steady-state and inflammatory dendritic-cell development. Nature Rev. Immunol. 7, 19–30 (2007).
Macatonia, S.E., Knight, S.C., Edwards, A.J., Griffiths, S. & Fryer, P. Localization of antigen on lymph node dendritic cells after exposure to the contact sensitizer fluorescein isothiocyanate. Functional and morphological studies. J. Exp. Med. 166, 1654–1667 (1987).
Leon, B., Lopez-Bravo, M. & Ardavin, C. Monocyte-derived dendritic cells formed at the infection site control the induction of protective T helper 1 responses against Leishmania. Immunity 26, 519–531 (2007).
Smith, C.M. et al. Cutting edge: conventional CD8α+ dendritic cells are preferentially involved in CTL priming after footpad infection with herpes simplex virus-1. J. Immunol. 170, 4437–4440 (2003).
Allan, R.S. et al. Epidermal viral immunity induced by CD8α+ dendritic cells but not by Langerhans cells. Science 301, 1925–1928 (2003).
Belz, G.T., Shortman, K., Bevan, M.J. & Heath, W.R. CD8α+ dendritic cells selectively present MHC class I–restricted noncytolytic viral and intracellular bacterial antigens in vivo. J. Immunol. 175, 196–200 (2005).
Belz, G.T. et al. Cutting edge: conventional CD8α+ dendritic cells are generally involved in priming CTL immunity to viruses. J. Immunol. 172, 1996–2000 (2004).
Belz, G.T. et al. Distinct migrating and nonmigrating dendritic cell populations are involved in MHC class I-restricted antigen presentation after lung infection with virus. Proc. Natl. Acad. Sci. USA 101, 8670–8675 (2004).
Allan, R.S. et al. Migratory dendritic cells transfer antigen to a lymph node–resident dendritic cell population for efficient CTL priming. Immunity 25, 153–162 (2006).
Carbone, F.R., Belz, G.T. & Heath, W.R. Transfer of antigen between migrating and lymph node–resident DCs in peripheral T-cell tolerance and immunity. Trends Immunol. 25, 655–658 (2004).
Croft, M., Bradley, L.M. & Swain, S.L. Naive versus memory CD4 T cell response to antigen. Memory cells are less dependent on accessory cell costimulation and can respond to many antigen-presenting cell types including resting B cells. J. Immunol. 152, 2675–2685 (1994).
Byrne, J.A., Butler, J.L. & Cooper, M.D. Differential activation requirements for virgin and memory T cells. J. Immunol. 141, 3249–3257 (1988).
Weninger, W., Crowley, M.A., Manjunath, N. & von Andrian, U.H. Migratory properties of naive, effector, and memory CD8+ T cells. J. Exp. Med. 194, 953–966 (2001).
Blaney, J.E., Jr. et al. Immunization with a single major histocompatibility complex class I–restricted cytotoxic T-lymphocyte recognition epitope of herpes simplex virus type 2 confers protective immunity. J. Virol. 72, 9567–9574 (1998).
Klenerman, P. & Zinkernagel, R.M. Original antigenic sin impairs cytotoxic T lymphocyte responses to viruses bearing variant epitopes. Nature 394, 482–485 (1998).
Mongkolsapaya, J. et al. Original antigenic sin and apoptosis in the pathogenesis of dengue hemorrhagic fever. Nat. Med. 9, 921–927 (2003).
Turner, S.J., Cross, R., Xie, W. & Doherty, P.C. Concurrent naive and memory CD8+ T cell responses to an influenza A virus. J. Immunol. 167, 2753–2758 (2001).
Soares, H. et al. A subset of dendritic cells induces CD4+ T cells to produce IFN-γ by an IL-12–independent but CD70-dependent mechanism in vivo. J. Exp. Med. 204, 1095–1106 (2007).
Tian, T., Woodworth, J., Skold, M. & Behar, S.M. In vivo depletion of CD11c+ cells delays the CD4+ T cell response to Mycobacterium tuberculosis and exacerbates the outcome of infection. J. Immunol. 175, 3268–3272 (2005).
Dewalick, S. et al. Cutting edge: conventional dendritic cells are the critical APC required for the induction of experimental cerebral malaria. J. Immunol. 178, 6033–6037 (2007).
Fleeton, M.N. et al. Peyer's patch dendritic cells process viral antigen from apoptotic epithelial cells in the intestine of reovirus-infected mice. J. Exp. Med. 200, 235–245 (2004).
He, Y., Zhang, J., Donahue, C. & Falo, L.D., Jr. Skin-derived dendritic cells induce potent CD8+ T cell immunity in recombinant lentivector-mediated genetic immunization. Immunity 24, 643–656 (2006).
Phillips, R.E. et al. Human immunodeficiency virus genetic variation that can escape cytotoxic T cell recognition. Nature 354, 453–459 (1991).
Pircher, H. et al. Viral escape by selection of cytotoxic T cell–resistant virus variants in vivo. Nature 346, 629–633 (1990).
Weiner, A. et al. Persistent hepatitis C virus infection in a chimpanzee is associated with emergence of a cytotoxic T lymphocyte escape variant. Proc. Natl. Acad. Sci. USA 92, 2755–2759 (1995).
Voeten, J.T. et al. Antigenic drift in the influenza A virus (H3N2) nucleoprotein and escape from recognition by cytotoxic T lymphocytes. J. Virol. 74, 6800–6807 (2000).
Mueller, S.N., Heath, W.R., Carbone, F.R. & Jones, C.M. The characterisation of two transgenic mice specific for herpes simplex virus. Immunol. Cell Biol. 80, 156–163 (2002).
Smith, C.M. et al. Cognate CD4+ T cell licensing of dendritic cells in CD8+ T cell immunity. Nat. Immunol. 5, 1143–1148 (2004).
Belz, G.T. et al. The CD8α+ dendritic cell is responsible for inducing peripheral self-tolerance to tissue-associated antigens. J. Exp. Med. 196, 1099–1104 (2002).
Manjunath, N. et al. Effector differentiation is not prerequisite for generation of memory cytotoxic T lymphocytes. J. Clin. Invest. 108, 871–878 (2001).
Klebanoff, C.A. et al. IL-15 enhances the in vivo antitumor activity of tumor-reactive CD8+ T cells. Proc. Natl. Acad. Sci. USA 101, 1969–1974 (2004).
Klebanoff, C.A. et al. Central memory self/tumor-reactive CD8+ T cells confer superior antitumor immunity compared with effector memory T cells. Proc. Natl. Acad. Sci. USA 102, 9571–9576 (2005).
Wong, P. & Pamer, E.G. Feedback regulation of pathogen-specific T cell priming. Immunity 18, 499–511 (2003).
Acknowledgements
We thank J. Langley, M. Camilleri and the WEHI Flow Cytometry Facility for technical assistance; and J. Miller for discussions. Supported by the National Health and Medical Research Council (Australia), the Wellcome Trust (G.T.B.), the Howard Hughes Medical Institute (G.T.B. and W.R.H.) and the Deutsche Forschungsgemeinschaft (BE 3285/1-1 and BE 3285/1-2 to S.B.).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–6 (PDF 323 kb)
Rights and permissions
About this article
Cite this article
Belz, G., Bedoui, S., Kupresanin, F. et al. Minimal activation of memory CD8+ T cell by tissue-derived dendritic cells favors the stimulation of naive CD8+ T cells. Nat Immunol 8, 1060–1066 (2007). https://doi.org/10.1038/ni1505
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ni1505
This article is cited by
-
Dendritic cell subsets in T cell programming: location dictates function
Nature Reviews Immunology (2019)
-
Memory responses of innate lymphocytes and parallels with T cells
Seminars in Immunopathology (2018)
-
The respiratory DC/macrophage network at steady-state and upon influenza infection in the swine biomedical model
Mucosal Immunology (2016)
-
Moving to the suburbs: T‐cell positioning within lymph nodes during activation and memory
Immunology & Cell Biology (2015)
-
The use of a TLR2 agonist‐based adjuvant for enhancing effector and memory CD8 T‐cell responses
Immunology & Cell Biology (2014)
