
Abstract
Excessive accumulation of white adipose tissue (WAT) is the defining characteristic of obesity. WAT mass is composed primarily of mature adipocytes, which are generated through the proliferation and differentiation of adipocyte precursors (APs). Although the production of new adipocytes contributes to WAT growth in obesity, little is known about the cellular and molecular mechanisms underlying adipogenesis in vivo. Here, we show that high-fat diet feeding in mice rapidly and transiently induces proliferation of APs within WAT to produce new adipocytes. Importantly, the activation of adipogenesis is specific to the perigonadal visceral depot in male mice, consistent with the patterns of obesogenic WAT growth observed in humans. Furthermore, we find that in multiple models of obesity, the activation of APs is dependent on the phosphoinositide 3-kinase (PI3K)-AKT2 pathway; however, the development of WAT does not require AKT2. These data indicate that developmental and obesogenic adipogenesis are regulated through distinct molecular mechanisms.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others

References
Kelly, T., Yang, W., Chen, C. S., Reynolds, K. & He, J. Global burden of obesity in 2005 and projections to 2030. Int. J. Obes. (Lond). 32, 1431–1437 (2008).
Peckham, S. C., Entenman, C. & Carroll, H. W. The influence of a hypercaloric diet on gross body and adipose tissue composition in the rat. J. Nutr. 77, 187–197 (1962).
Steinberg, M. D., Zingg, W. & Angel, A. Studies of the number and volume of fat cells in adipose tissue. J. Pediatr. 61, 299–300 (1962).
Enesco, M. & Leblond, C. P. Increase in cell number as a factor in the growth of the organs and tissues of the young male rat. J. Embryol. Exp. Morphol. 10, 530–562 (1962).
Spalding, K. L. et al. Dynamics of fat cell turnover in humans. Nature 453, 783–787 (2008).
Arner, P. et al. Variations in the size of the major omentum are primarily determined by fat cell number. J. Clin. Endocrinol. Metab. 98, E897–E901 (2013).
Phillips, L. K. & Prins, J. B. The link between abdominal obesity and the metabolic syndrome. Curr. Hypertens. Rep. 10, 156–164 (2008).
Bjorntorp, P. Metabolic difference between visceral fat and subcutaneous abdominal fat. Diabetes Metab. 26 (Suppl. 3), 10–12 (2000)
Wang, Y., Rimm, E. B., Stampfer, M. J., Willett, W. C. & Hu, F. B. Comparison of abdominal adiposity and overall obesity in predicting risk of type 2 diabetes among men. Am. J. Clin. Nutr. 81, 555–563 (2005).
Hirsch, J. & Batchelor, B. Adipose tissue cellularity in human obesity. Clin. Endocrinol. Metab. 5, 299–311 (1976).
Faust, I. M., Johnson, P. R., Stern, J. S. & Hirsch, J. Diet-induced adipocyte number increase in adult rats: a new model of obesity. Am. J. Physiol. 235, E279–E286 (1978).
Lemonnier, D. Effect of age, sex, and sites on the cellularity of the adipose tissue in mice and rats rendered obese by a high-fat diet. J. Clin. Invest. 51, 2907–2915 (1972).
Wang, Q. A., Tao, C., Gupta, R. K. & Scherer, P. E. Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat. Med. 19, 1338–1344 (2013).
Cleary, M. P., Brasel, J. A. & Greenwood, M. R. Developmental changes in thymidine kinase, DNA, and fat cellularity in Zucker rats. Am. J. Physiol. 236, E508–E513 (1979).
Cristancho, A. G. & Lazar, M. A. Forming functional fat: a growing understanding of adipocyte differentiation. Nat. Rev. Mol. Cell Biol. 12, 722–734 (2011).
Rodeheffer, M. S., Birsoy, K. & Friedman, J. M. Identification of white adipocyte progenitor cells in vivo. Cell 135, 240–249 (2008).
Jeffery, E. et al. Characterization of Cre recombinase models for the study of adipose tissue. Adipocyte 3, 206–211 (2014).
Muzumdar, M. D., Tasic, B., Miyamichi, K., Li, L. & Luo, L. A global double-fluorescent Cre reporter mouse. Genesis 45, 593–605 (2007).
Berry, R. & Rodeheffer, M. S. Characterization of the adipocyte cellular lineage in vivo. Nat. Cell Biol. 15, 302–308 (2013).
Pellettieri, J. & Sanchez Alvarado, A. Cell turnover and adult tissue homeostasis: from humans to planarians. Annu. Rev. Genet. 41, 83–105 (2007).
Joe, A. W., Yi, L., Even, Y., Vogl, A. W. & Rossi, F. M. Depot-specific differences in adipogenic progenitor abundance and proliferative response to high-fat diet. Stem Cells 27, 2563–2570 (2009).
Hudak, C. S. et al. Pref-1 marks very early mesenchymal precursors required for adipose tissue development and expansion. Cell Rep. 8, 678–687 (2014).
Church, C. D., Berry, R. & Rodeheffer, M. S. Isolation and study of adipocyte precursors. Methods Enzymol. 537, 31–46 (2014).
Berry, R., Jeffery, E. & Rodeheffer, M. S. Weighing in on adipocyte precursors. Cell Metab. 19, 8–20 (2014).
Lee, Y. H., Petkova, A. P., Mottillo, E. P. & Granneman, J. G. In vivo identification of bipotential adipocyte progenitors recruited by β3-adrenoceptor activation and high-fat feeding. Cell Metab. 15, 480–491 (2012).
Hietakangas, V. & Cohen, S. M. Regulation of tissue growth through nutrient sensing. Annu. Rev. Genet. 43, 389–410 (2009).
Pearce, L. R., Komander, D. & Alessi, D. R. The nuts and bolts of AGC protein kinases. Nat. Rev. Mol. Cell Biol. 11, 9–22 (2010).
Eto, H. et al. Characterization of structure and cellular components of aspirated and excised adipose tissue. Plast. Reconstr. Surg. 124, 1087–1097 (2009).
Gonzalez, E. & McGraw, T. E. The Akt kinases: isoform specificity in metabolism and cancer. Cell Cycle 8, 2502–2508 (2009).
Cho, H., Thorvaldsen, J. L., Chu, Q., Feng, F. & Birnbaum, M. J. Akt1/PKBα is required for normal growth but dispensable for maintenance of glucose homeostasis in mice. J. Biol. Chem. 276, 38349–38352 (2001).
Chen, W. S. et al. Growth retardation and increased apoptosis in mice with homozygous disruption of the Akt1 gene. Genes Dev. 15, 2203–2208 (2001).
Cho, H. et al. Insulin resistance and a diabetes mellitus-like syndrome in mice lacking the protein kinase Akt2 (PKB β). Science 292, 1728–1731 (2001).
Garofalo, R. S. et al. Severe diabetes, age-dependent loss of adipose tissue, and mild growth deficiency in mice lacking Akt2/PKB β. J. Clin. Invest. 112, 197–208 (2003).
Gautron, L. & Elmquist, J. K. Sixteen years and counting: an update on leptin in energy balance. J. Clin. Invest. 121, 2087–2093 (2011).
Han, J. et al. The spatiotemporal development of adipose tissue. Development 138, 5027–5037 (2011).
Birsoy, K. et al. Analysis of gene networks in white adipose tissue development reveals a role for ETS2 in adipogenesis. Development 138, 4709–4719 (2011).
Pollack, A. A.M.A. Recognizes Obesity as a Disease New York Times Online (18 June 2013); http://www.nytimes.com/2013/06/19/business/ama-recognizes-obesity-as-a-disease.html
Leavens, K. F., Easton, R. M., Shulman, G. I., Previs, S. F. & Birnbaum, M. J. Akt2 is required for hepatic lipid accumulation in models of insulin resistance. Cell Metab. 10, 405–418 (2009).
Czech, M. P., Tencerova, M., Pedersen, D. J. & Aouadi, M. Insulin signalling mechanisms for triacylglycerol storage. Diabetologia 56, 949–964 (2013).
Tontonoz, P. & Spiegelman, B. M. Fat and beyond: the diverse biology of PPARγ. Annu. Rev. Biochem. 77, 289–312 (2008).
Lodhi, I. J. et al. Inhibiting adipose tissue lipogenesis reprograms thermogenesis and PPARγ activation to decrease diet-induced obesity. Cell Metab. 16, 189–201 (2012).
George, S. et al. A family with severe insulin resistance and diabetes due to a mutation in AKT2. Science 304, 1325–1328 (2004).
Hussain, K. et al. An activating mutation of AKT2 and human hypoglycemia. Science 334, 474 (2011).
Tran, T. T., Yamamoto, Y., Gesta, S. & Kahn, C. R. Beneficial effects of subcutaneous fat transplantation on metabolism. Cell Metab. 7, 410–420 (2008).
Wajchenberg, B. L. Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocr. Rev. 21, 697–738 (2000).
Yamamoto, Y. et al. Adipose depots possess unique developmental gene signatures. Obesity 18, 872–878 (2010).
Grove, K. L., Fried, S. K., Greenberg, A. S., Xiao, X. Q. & Clegg, D. J. A microarray analysis of sexual dimorphism of adipose tissues in high-fat-diet-induced obese mice. Int. J. Obes. (Lond.) 34, 989–1000 (2010).
Macotela, Y., Boucher, J., Tran, T. T. & Kahn, C. R. Sex and depot differences in adipocyte insulin sensitivity and glucose metabolism. Diabetes 58, 803–812 (2009).
Chau, Y. Y. et al. Visceral and subcutaneous fat have different origins and evidence supports a mesothelial source. Nat. Cell Biol. 16, 367–375 (2014).
Krueger, K. C., Costa, M. J., Du, H. & Feldman, B. J. Characterization of cre recombinase activity for in vivo targeting of adipocyte precursor cells. Stem Cell Rep. 3, 1147–1158 (2014).
Rosen, E. D. & Spiegelman, B. M. What we talk about when we talk about fat. Cell 156, 20–44 (2014).
Van Herpen, N. A. & Schrauwen-Hinderling, V. B. Lipid accumulation in non-adipose tissue and lipotoxicity. Physiol. Behav. 94, 231–241 (2008).
Berry, R. et al. Imaging of adipose tissue. Methods Enzymol. 537, 47–73 (2014).
Acknowledgements
This work was supported by American Diabetes Association Award 7-12-JF-46, DERC pilot project grant DK045735 and NIDDK grant DK090489 to M.S.R., Lo Graduate Fellowship for Excellence in Stem Cell Research from the Yale Stem Cell Center to E.J., and EMBO long-term fellowship ALTF 132-2011 to C.D.C.
Author information
Authors and Affiliations
Contributions
E.J., C.D.C. and M.S.R. designed experiments. E.J., C.D.C., B.H., L.C. and M.S.R. performed experiments. E.J., C.D.C. and B.H. analysed data. E.J., C.D.C., B.H. and M.S.R. interpreted data. E.J. and M.S.R. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 3 Characterization of HFD-induced adipocyte hyperplasia.
(A) Experimental scheme for BrdU time course in Figure 2B. (B) Representative flow cytometry dot plots indicating sequential gating used to identify AP populations, including CD24+ adipocyte progenitors (CD45−, CD31−, CD34+, CD29+, Sca1+, CD24+), CD24− preadipocytes (CD45−, CD31−, CD34+, CD29+, Sca1+, CD24−), and total APs (CD45−, CD31−, CD34+, CD29+, Sca1+). (C) Flow cytometry plots of SVF from VWAT of PdgfRα-H2B-GFP mice showing that the majority of GFP+ cells are CD29+; CD34+, of which 97.1 ± 1.5% are also Sca1+. (n = 3 mice) (D) Quantification of BrdU incorporation into GFP+ cells from VWAT of male PdgfRα-H2B-GFP mice during 1 week of SD or HFD feeding and BrdU treatment. (n = 5 mice for SD and 7 mice for HFD.) Significance between the indicated groups in (D) was calculated using a two-tailed Student’s t-test. Exact p-values are listed in Supplementary Table 1. Error bars or values represent mean ± s.e.m.∗∗ (p < 0.01). HFD: high-fat diet, SD: standard diet, BrdU: bromodeoxyuridine.
Supplementary Figure 4 Adipocyte precursor proliferation during week 2 of HFD feeding does not lead to adipogenesis.
(A) Schematic and depicting the second week BrdU pulse-chase in Supplementary Figure 2B. (B) BrdU incorporation into adipocyte nuclei after pulse-chase from the second week of HFD feeding. (n = 5 for all groups). Error bars or values represent mean ± s.e.m. HFD: high-fat diet, SD: standard diet, BrdU: bromodeoxyuridine.
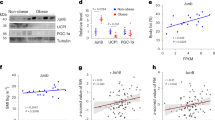
Supplementary Figure 5 High-fat diet does not induce AKT phosphorylation in whole fat.
(A) Western blot analysis for phosphorylated AKT at the indicated sites and total AKT, in lysates from whole VWAT on day 3 of diet. Each lane represents one animal. Uncropped blots are shown in Supplementary Figure 7I-L. (B) Quantification of pAKT western blots in (A), normalized to total AKT. (C-D) VWAT was treated with the indicated concentrations of insulin prior to tissue digestion, and APs were isolated in the presence of wortmannin and phosphatase inhibitors to limit further changes in pathway activation (see methods). APs were then fixed and analyzed by flow cytometry for levels of phosphorylated AKT at threonine 308 (C) or serine 473 (D). The same AP cell isolation procedure was used in the experiments in Figure 4B-F and 5B-C. (E) Western blot for phosphorylated AKT1 (S473) with lysates from APs enriched from SVF via Sca-1 bead pull down (Sca1-positive) and remaining unselected cells (Sca1-negative) (see methods) after 3 days of HFD or SD. Each lane represents pooled cells from 2 mice. Uncropped blots are shown in Supplementary Figure 7M-N. Significance between the indicated groups in (B) was calculated using a two-tailed Student’s t-test. Error bars represent mean ± s.e.m. HFD: high-fat diet, SD: standard diet.
Supplementary Figure 6 Deletion of AKT2 in the adipocyte lineage does not affect body weight or food intake.
(A) Quantification of BrdU incorporation into SWAT APs after 1 week of HFD or SD with daily injection of wortmannin (Wort), or vehicle (Veh) (n = 5 mice for each group). (B) PCR analysis of DNA isolated from AP cells from the VWAT and SWAT depots of female mice show successful excision of the floxed exons 4 and 5 from Akt2 gene, while Lin+ cells (CD45+ blood cells and CD31+ endothelial cells) contain the full-length Akt2 gene. Each lane represents cells isolated from the indicated depot of individual mice. (C) Body weight of Akt2fx/fl (n = 9 mice) and PdgfRα-cre; Akt2fl/fl (n = 12 mice) at 6 weeks of age. (D) Quantification of food intake normalized to body weight in the indicated groups of mice on SD or during the first 4 days of HFD feeding. (n = 3 mice for Akt2fl/fl SD and PdgfRα-cre; Akt2fl/flSD, n = 4 mice for PdgfRα-cre HFD, n = 2 mice for Akt2fl/fl HFD, and n = 3 mice for PdgfRα-cre; Akt2fl/fl HFD. (E) BrdU incorporation into SWAT APs of the indicated groups during the first week of HFD feeding compared to SD controls. (n = 7 mice for Akt2fl/fl SD, n = 4 mice for PdgfRα-cre; Akt2fl/fl SD, n = 14 mice for PdgfRα-cre; Akt2fl/fl HFD, n = 8 mice for Akt2fl/fl HFD, n = 5 mice for wild-type and PdgfRα-cre HFD). Significance in (D) was calculated using one-way ANOVA with Tukey’s test for multiple comparisons. Exact p-values are listed in Supplementary Table 1. Statistics source data for 4D can be found in Supplementary Table 2. Error bars represent mean ± s.e.m.∗∗ (p < 0.01),∗∗∗∗ (P < 0.0001).
Supplementary Figure 7 AKT2 is required for the activation of adipocyte precursors at the onset of obesity in ob/ob mice.
(A-B) Weekly intake of SD in raw kilocalories (kCal) (A) or kCal normalized to body weight (B) for the indicated groups of young mice after weaning at 3 weeks of age. Week “3” denotes food intake between the ages of 3 and 4 weeks. (n = 4 for wild-type, n = 3 for ob/ob, n = 7-8 for ob/ob; Akt2−/−) (C-D) H&E-stained sections of VWAT (C) and corresponding adipocyte size measurements (D) for the indicated groups. (n = 7 for ob/ob; Akt2−/−, n = 4 for ob/ob). Significance between indicated groups in (A-B) was calculated using a two-tailed Student’s t-test. Significance in (D) was calculated using two-way ANOVA with Bonferroni’s test for multiple comparisons. Scale bars in (C) are 50 μm.∗ or a (P < 0.05),∗∗ or b (p < 0.01),∗∗∗ or c (P < 0.001),∗∗∗∗ or d (P < 0.0001). For (A-B), ∗ indicates significance of ob/ob over wild-type, † indicates significance of ob/ob; Akt2−/− over wild-type, and ‡ indicates significance of ob/ob; Akt2−/− over ob/ob. Exact p-values are listed in Supplementary Table 1. Statistics source data for 5A-B can be found in Supplementary Table 2. Error bars represent mean ± s.e.m.
Supplementary Figure 8 AKT2 is not required for the normal development of white adipose tissue.
(A-B) Confocal images (A) of developing wild-type and Akt2−/− SWAT stained with LipidTOX (a lipid stain) and Isolectin IB4 (an endothelial cell stain) and (B) corresponding lipid droplet size quantification based on LipidTOX staining (n = 3 mice for all groups except n = 4 mice for Akt2−/−P5). Each group includes mice from 2-3 different litters. (C) Echo MRI quantification of fat mass from the indicated groups of mice at 6 weeks of age. (n = 13 mice for Akt2fl/fl and n = 14 mice for PdgfRα-cre; Akt2fl/fl). Scale bars in (A) are 50 μm. Error bars represent mean ± s.e.m. PX indicates postnatal day X.
Supplementary information
Supplementary Information
Supplementary Information (PDF 592 kb)
Rights and permissions
About this article

Cite this article
Jeffery, E., Church, C., Holtrup, B. et al. Rapid depot-specific activation of adipocyte precursor cells at the onset of obesity. Nat Cell Biol 17, 376–385 (2015). https://doi.org/10.1038/ncb3122
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ncb3122
This article is cited by
-
Single-cell analysis reveals a subpopulation of adipose progenitor cells that impairs glucose homeostasis
Nature Communications (2024)
-
Maternal obesity and programming of metabolic syndrome in the offspring: searching for mechanisms in the adipocyte progenitor pool
BMC Medicine (2023)
-
Adipokines at the crossroads of obesity and mesenchymal stem cell therapy
Experimental & Molecular Medicine (2023)
-
Leptin receptor+ cells promote bone marrow innervation and regeneration by synthesizing nerve growth factor
Nature Cell Biology (2023)
-
Autologous adipose-derived stromal vascular fraction and platelet concentrates for the treatment of complex perianal fistulas
Techniques in Coloproctology (2023)
