
Abstract
Myotonia congenita is a genetic disease characterized by impaired muscle relaxation after forceful contraction (myotonia) and caused by mutations in the chloride channel voltage-sensitive 1 (CLCN1) gene, encoding the voltage-gated chloride channel of skeletal muscle (ClC-1). In a large cohort of clinically diagnosed unrelated probands, we identified 75 different CLCN1 mutations in 106 individuals, among which 29 were novel mutations and 46 had already been reported. Despite the newly described mutations being scattered throughout the gene, in our patients, mutations were mostly found in exons 4 and 5. Most of the novel mutations located in the region comprising the intramembrane helices are involved in the ion-conducting pathway and predicted to affect channel function. We report for the first time that two mutations, inherited on the same allele as a heterozygous trait, abrogate disease expression, although when inherited singularly they were pathogenic. Such a mode of inheritance might explain the incomplete penetrance reported for autosomal dominant mutations in particular families.
Similar content being viewed by others


Introduction
Myotonia, an abnormal delay in muscle relaxation following voluntary forceful contraction, more pronounced after inactivity, is the prominent symptom of myotonia congenita (MC).1, 2, 3 Affected individuals report muscular stiffness upon initiating movement, which remits with several repetitions of the same movement (‘warm-up’ phenomenon).3 MC may be transmitted either as an autosomal dominant (Thomsen disease, OMIM #160800) or a recessive trait (Becker disease, OMIM #255700).4 Although Thomsen2 and Becker1 did not find any sex-related differences in MC frequency, a significant difference, related to gender, is reported in recessive, but not in dominant, MC, men being more severely affected than women. Both forms of MC are due to mutations in the chloride channel voltage-sensitive 1 (CLCN1) gene (OMIM*118425), located on chromosome 7q35 and organized into 23 exons.5 The gene encodes the main human skeletal muscle chloride channel (ClC-1), important for the normal repolarization of muscle action potential. The X-ray crystallography study of the prokaryotic ClC Cl− channels reveals two identical subunits that form a homodimeric membrane protein; each subunit forms its own ion-conduction pore and exhibits an antiparallel architecture that defines a selectivity filter for the Cl anion.6 The subunits contain 18 α-helices (labeled A–R) of which 16 (B–Q) are transmembrane. All ClC proteins have a long cytoplasmic carboxy terminus that contains two identical cystathionine b-synthase (CBS) domains that interact; it has been proposed that the CBS domains have a functional role in the common gate that closes the pores of the double-pore channel simultaneously.7 Loss of function of the chloride channel renders the plasma membrane hyper-excitable, leading to the typical ‘myotonic runs’ seen in electromyograms of myotonic patients.8
More than 130 different CLCN1 mutations, scattered throughout the gene, have been described to be associated with MC; they include deletions, insertions, frame-shifts, stop codons, missense and splice-site mutations.3, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 Most of these mutations are recessive and in the majority of cases occur as compound heterozygous mutations, albeit not always demonstrated biochemically.3 The same mutations can be associated with either dominant or recessive pedigrees, indicating an incomplete penetrance or variable expressivity of certain dominant mutations.3, 8, 11, 15, 20 Only a few published mutations have been demonstrated to be dominant, through electrophysiological assessment.13, 15 A common phenotype of dominant mutations is a dominant negative effect of mutant subunits in mutant–wild-type heterodimers, causing a large shift of the steady-state open probability voltage-dependence towards more positive, non-physiological voltages.15
The purpose of this study was to characterize the entire coding region of the CLCN1 gene in a large cohort of Italian patients to define genetic epidemiology and identify population-specific genetic features. Bioinformatics mutation analysis, which models structural/functional effect between wild-type and mutant amino acid, was applied for studying the effect of novel missense and intronic mutations, comparing also the identified novel mutations with those located in the same region of the protein and already characterized for their electrophysiological properties.
MATERIALS AND METHODS
Patients
Three hundred and forty-five subjects were studied: 215 probands (67% male (143/215) and 33% female (72/215) individuals; age range, 5–72 years), included on the basis of clinical myotonic symptoms and/or EMG-confirmed myotonic recordings,21 and their 130 relatives. Of the 215 probands, 197 were of Italian origin and 18 were of other ethnic groups. Myotonic dystrophy type 1 was excluded in all patients by genetic screening. Written informed consent for DNA storage and use for genetic analysis and research purposes were obtained from all patients (parents or tutors for patients under age) and relatives, as required by the Ethical Committee of Foundation Neurological Institute Carlo Besta.
Genetic screening by PCR amplification and sequence analysis of CLCN1 exons
Genomic DNA was extracted from peripheral blood lymphocytes by standard procedures and screened for CLCN1 mutations by PCR analysis of the 23 exons, as previously described by Brugnoni et al.9 The PCR products were purified by ExoSAP-IT (USB-Affymetrix, Cleveland, OH, USA) and sequenced by bidirectional sequencing using the BigDye Terminator v3.1 Cycle Sequencing Kit (Life Technologies, Foster City, CA, USA), on an ABI PRISM 3100 Genetic Analyzer (Life Technologies). The obtained sequences were analyzed with SeqScape v.2.1.1 software (Life Technologies) and compared with reference wild-type sequence (GenBank accession numbers: CAA81103.1; NM_000083.2).
All novel mutations were confirmed by sequence analysis of a second independent PCR product and by screening 100 unrelated healthy Italian individuals who gave their informed consent for using their DNA for genetic research, as controls (200 alleles). The variants have been submitted to the Leiden Open Variation Database (http://chromium.liacs.nl/LOVD2/).19
Bioinformatics analysis: prediction of the three-dimensional structure of ClC-1 by homology modelling
The human ClC-1 amino acid sequence was obtained from RefSeq database (NP_000074.2) to build two homology models: transmembrane (TrClC-1) and C-terminal CBS regions (CtClC-1) using X-ray crystallographic templates (PDB id: 1KPL and 24AZ, respectively) obtained from Protein Data Bank (www.rcsb.org). Homology modeling of the ClC-1 was performed using MODELLER.22 The two models in construction shared 26% identity and 30% similarity for TrClC-1 (F161 to S572) and 54% identity and 58% similarity for CtClC-1 (M561 to A862). The linker between CBS1 and CBS2 domains does not share high homology with ClC-1 residues 669–820 and, therefore, was excluded from CtClC-1. Energy minimization was applied using GROMACS 4 force field.23 Stereochemistry was checked after energy minimization using PROCHECK.24 The PROCHECK results for the TrClC-1 and CtClC-1 models showed >96% of residues in allowed regions of the Ramachandran plot. The localization of mutations was visualized using the PyMOL software (http://www.pymol.org).
The MutPred software (http://mutpred.mutdb.org/) was used to predict if a mutation can have deleterious effects and what is its possible molecular consequence on the protein. Splice-site variations for modifications of exonic splicing enhancers or splicing acceptor-donor sites were evaluated using Human Splicing Finder v.2.4.1 (http://www.umd.be/HSF/).25, 26 The evolutionary conservation score among the ClC-1 protein homologs was calculated by ConSurf server using the Bayesian method (http://consurf.tau.ac.il/).
Results
Distribution of CLCN1 mutations in the MC cohort
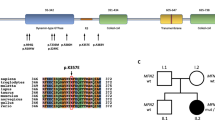
CLCN1 gene mutations were detected in 106 (49.3%) of the 215 probands (69% male (73/106) and 31% female subjects (33/106)), 93 of these were of Italian origin, and 68 (52.3%) of the 130 relatives. In the 106 probands, we found 75 different mutations scattered across the 23 exons of the CLCN1 gene: 46 (61.3%) known and 29 (38.7%) novel (Supplementary Table 1). Those unreported included 20 missense, 4 nonsense, 2 splice-site and 3 deletion mutations (Supplementary Table 1 and Figure 1a). None of the novel mutations were detected in 200 control chromosomes. The novel mutations were detected in single patients or families, except for the following three mutations: p.R338X was the most frequent, as it was found in four probands (one of whom was of Iranian origin); p.A535D and p.V536L were each found in two probands (Supplementary Table 1 and Figure 1a).

Distribution of the 156 novel and known CLCN1 mutations detected in the 106 probands. (a) Novel nucleotide mutations; the amino-acid variants are indicated in parentheses. (b) Known nucleotide mutations. A full color version of this figure is available at the Journal of Human Genetics journal online.
Among the 46 known CLCN1 mutations,9, 11, 12, 14, 15, 17, 18, 27, 28 which included 28 missense, 7 nonsense, 8 splice-site and 3 deletion mutations (Supplementary Table 1 and Figure 1b), three were very common: the splice-site mutation c.180+3A>T in intron 1, the missense mutation c.501C>G (p.F167L) in exon 4 and the missense mutation c.568-569GG>TC (p.G190S) in exon 5 identified in 11, 16 and 12 unrelated MC patients, respectively (Supplementary Table 1 and Figure 1b).21, 27, 29
Among the 106 CLCN1-mutated MC probands, we found 49 with one mutated allele and 57 with two mutated alleles. Among the 49 MC probands with one mutated allele, five had a dominant family history and, therefore, they are likely to be affected by a dominant form of MC (Thomsen disease); five carried a mutation reported in the literature inherited either in a dominant or in a recessive mode;9, 12 five harbored a mutation reported in the literature to be inherited in a recessive mode.9, 12 For the remaining 34 cases, the presence of a second, yet undetected, mutation cannot be excluded and therefore firm conclusions regarding the inheritance of mutation require functional experiments and the study of other family members carrying the same mutation. Thus, 63 of the 106 probands harbored a single heterozygous (n=49) or homozygous (n=14) mutation, 40 probands carried two different mutations, and 3 probands presented with three heterozygous mutations. The three mutations were observed in one Italian family (p.R105C, p.F167L and p.Q812X: described below) and in two probands of African origin (p.R105C, p.F167L, p.V217D and p.F167L, p.R421C, p.P932L, respectively).
Predicted effect of the novel mutations
Effects of the novel mutations that do not result in a stop codon, deletion or splice mutation were predicted by MutPred and are summarized in Table 1. Modeling of the structural model of the ClC-1 (Supplementary Figure 1) reveals that, with the exception of mutations p.R453W, p.V217D and p.G395E, most of the missense mutations identified in this study reside within or in close proximity to the Cl− selectivity filter. Two novel mutations transformed the first amino acid of a transmembrane α-helix from proline to arginine (p.P282R and p.P521R), where rigidity of the proline residue might be essential to maintain the spatial conformation of the domain (Table 1 and Supplementary Figure 1b).
Interestingly, the two splicing sites, c.2364+10G>A and c.2364+5G>A, analyzed by Human Splice Finder (HSF),25, 26 showed that they are implicated in the abrogation and creation of enhancers/silencer-binding sites as well as of splice acceptor, and thereby might have an impact on splicing (Table 2).
Multiple sequence alignment of 108 unique ClC-1 homologs show that 55% (549/989) of residues are evolutionarily conserved (conservation score ⩾6). Of these conserved residues, 167 residues are very highly evolutionarily conserved (score=9) and are expected to be in functionally important segments of the ClC-1. In our study, seven identified mutations concerned highly conserved residues (p.G188A, p.V273M, p.P282R, p.P521R, p.V536L, p.V640G and p.L520P). Of the remaining 19 missense mutations analyzed in this study, 8 (p.L198P, p.V217D, p.A244S, p.R338X, p.G395E, p.F484L, p.F494S and p.A535D) were also conserved residues (6Table 1).
Effects of the novel mutations that resulted in a stop codon or deletion, which led to a truncated protein, are summarized in Table 3.
Discussion
Our genetic screening of 215 probands with suspected MC identified 106 mutated, unrelated probands. Among the different mutations detected, 29 were novel and 46 were previously reported.9, 12, 13, 14, 15, 17, 18, 27, 28 As shown in Figure 2, the 75 different mutations are scattered across the entire chloride channel, with the exception of the regions spanning α-helices M and Q and the cytoplasmic helix R, in which we did not detect any mutation. Like in other cohort studies of different ethnicities including Southern European, Northern European and French-Canadian individuals,9, 10, 13, 14, 16, 17, 30, 31, 32 there was no clear evidence of founder-effect manifestations for the 75 different CLCN1 mutations detected in our proband cohort.

Localization on the CLC-1 protein of the 75 mutations detected in this study. The scheme shows the putative structure of the subunit as predicted by Dutzler et al.6 and Estévez et al.7 The 29 novel mutations are labeled with the amino acid change and shown as red squares. The known mutations are shown as yellow squares, with the amino acid change shown in bold only for the three most frequently occurring, c.180+3A>T, p.F167L, and p.G190S.
Of the 20 novel missense mutations, which involve amino acids highly evolutionarily conserved, 7 were located within or in the close vicinity of the Cl– selectivity filter (except p.R453W and p.A244S), and therefore more likely to reduce directly binding affinity to Cl− (Supplementary Figure 1).
Our study identified a region of high mutation frequency that includes exons 4 and 5 of the CLCN1 gene: 37 (34.9%) unrelated probands harbored mutations in this region (Figure 1); other exons with a high rate of mutations, although to a less frequency, included exons 1, 11, 13, 15 and 23 (Figure 1). Out of the known mutations, three were more frequent in our cohort: the c.180+3A>T splice-site mutation and the two missense mutations p.F167L and p.G190S in exon 4 and 5, respectively.27, 33, 34
The c.180+3A>T mutation probably implies the deletion of exon 1 resulting in a non-functional gene product, and therefore in a lack of, or decrease in, functional chloride channels leading to MC with diverse phenotypes.21, 31 A recent report on the largest Spanish MC cohort investigated so far indicated that 50% of the Spanish families carried this splicing mutation, either in homozygous or compound heterozygous form, suggesting that such a mutation is highly frequent among the Spanish.13 This was confirmed by our data from the largest Italian cohort reported so far (93 of 106 mutated probands were of Italian origin): only 12% of our Italian probands carried the mutation c.180+3A>T (Table 4). In another study on Italian patients, albeit smaller (29 probands), the mutation was also observed at low frequency (10.3%),14 supporting the tight correlation of the mutation with a Spanish origin of the patients (Table 4). These frequencies are nevertheless higher than those observed in other European populations, albeit each also of considerably smaller size than our cohort (Table 4). According to the model of Duztler et al.,6 the F167 located in the α-helix C of ClC-1 (Figure 2) is conserved in all known ClC chloride channels;29 electrophysiological studies have shown that p.F167L mutation causes a slight shift of the open probability of the channel, thus altering normal muscle excitability.35
The third most frequent mutation, p.G190S, also occurs in a well-conserved region of the polypeptide loop that precedes the α-helix D of the channel (Figure 2).6 The p.G190S mutation was recently reported in a large consanguineous Arab family, where it occurred both in the heterozygous state in asymptomatic or mildly affected individuals and in the homozygous state in severely affected individuals.27 Interestingly, when comparing the reported European cohorts, this mutation only appears in the Italian cohorts (Table 4).
Another mutation only observed in the two Italian cohorts is p.I290M (Table 4). It is located in exon 8 that codes for a highly conserved region of predicted interaction between the ClC-1 monomers; based on data obtained from a large cohort of UK patients, exon 8 was proposed to be a hotspot for dominant mutations.12 In contrast, other mutations not detected or scarce in our and other Southern European cohorts, in particular p.F413C, were more prominent in Northern European countries (Table 4). Interestingly, p.R894X accounted for <10% in the European cohorts reported, including ours, except in the small Northern Scandinavian cohort where it was observed in 55.5% of probands (Table 4).
Eighteen of the novel mutations detected in our cohort were located within the region that includes transmembrane helices B–Q (Figure 2). Our novel dominant missense mutations do not cluster on exon 8 as proposed by Fialho et al.,12 therefore suggesting domains other than H and I helices to be involved in the molecular mechanism of the disease. Two of the dominant amino acid changes introduce a Pro residue in helix D (p.L198P) and at the end of helix N (p.L520P). In particular, helix D is part of the pore domain, hence, such a structural alteration may account for the elevated deleterious index calculated for the p.L198P mutant. Other novel dominant mutations include p.A244S and p.R453D located in the core of the protein complex. Two novel (p.G188A, p.F484L) and two known (p.G190S, p.G482R) detected mutations reside in the ion-conducting pathway and affect residues involved in ion selectivity and conductance. Consistent with their location, the deleterious index calculated for the two novel mutations is very high, predicting altered ion selectivity and disrupted function. Indeed, a cysteine-scanning mutagenesis study revealed that the substitution of G188 with a cysteine leads to a dramatic reduction in chloride current.36 Moreover, the p.G190S mutant, with a deleterious index of 0.83, has been recently shown to affect open probability, deactivation properties, permeability and current densities, whereas channels bearing the two pore mutations p.G230V and p.G233V, with a deleterious index of 0.965 and 0.961, were not functional at all.13, 37 Therefore, these functional evidences validate the correlation between the numerical prediction and the molecular defect for dominant and recessive mutations located in the channel pore.
Mutagenesis and electrophysiological studies revealed the importance of the cytoplasmic C-terminal structure, with the highly conserved interacting CBS domains, for ClC-1 functional expression, common gate modulation, nucleotide and protein kinase binding.7, 38, 39 Ten of the 29 novel mutations detected in our cohort were located within the cytoplasmic C-terminal region (Figure 2). Four of these were dominant (Tables 1 and 3) and are likely to affect the interaction of ClC-1 with other proteins or interactions that occur within ClC-1.7 The mutation c.2786delC (p.T929TfsX19, Table 3) results in a stop codon that truncates the last 22 amino acids of the protein and is expected to dramatically reduce channel expression and shift the voltage of half-maximal activation.7, 40 Indeed, the p.R894X mutant, which truncates the protein after the CBS2 domain, reduced current amplitude in parallel with its surface expression level.40 Three dominant mutations reside in the CBS1–CBS2 linker: the splice-site mutation c.2364+10G>A (Figure 2, Table 2) and the two missense mutations p.D687H and p.Q763K (Figure 2, Table 1). They may modify the C-terminal protein structure within the stretch between CBS1 and CBS2 and/or likely impair the interaction of the CBS2 domains with the membrane-resident portion of the protein, thus affecting common gating.7, 41 Bennetts et al.39 highlighted three residues on both CBS1 and CBS2 comprising the putative site for ATP binding. Cytoplasmic ATP has been shown to block chloride currents by inhibiting the common gating. Interestingly, among the novel recessive mutations, the p.V640G (Figure 2, Table 1) resides very close to the putative ATP binding sites on CBS1 domain and a disruption of ATP block may result from its high deleterious index.39 In our study, ClC-1 molecular modeling of CBS domains demonstrated that these mutations influence the local structure and charge distribution. Additionally, on the basis of in silico analysis, it seems that the splice mutations identified in our study may affect the splicing process (Table 2).
In our MC families, the molecular characteristics of two brothers with autosomal recessive myotonia, who harbor three different mutations (p.R105C/p.F167L/p.Q812X), were of particular interest (data not shown). Such an unusual finding was described only in single patients in two recent studies.13, 42 It is most interesting to note that the two mutations p.R105C and p.F167L were found in association with a third mutation in this familial case and in one of the two sporadic cases in our cohort, as well as in both patients reported to carry three mutations in the other two studies.13, 42 In our family with three mutations, the p.R105C/p.F167L double mutation was only associated with disease in the presence of the third mutation, p.Q812X. Indeed, the mother who does not carry the third mutation is asymptomatic. The mode of inheritance of p.R105C and p.F167L indicates that both mutations are on the same allele and become pathogenic when in combination with p.Q812X. As also evident from the inheritance of p.Q812X in another family, this mutation is pathogenic both in homozygous and compound heterozygous modes (data not shown). Both p.R105C and p.F167L can be pathogenic on their own in heterozygous mode,34 but we did not detect p.R105C other than in combination with p.F167L in our cohort where, when both mutations were inherited in the homozygous mode in one proband, the combination was pathogenic (data not shown). It would therefore seem that when p.R105C and p.F167L are carried on the same allele in the heterozygous mode, the combination abrogates the pathogenic effect of each mutation taken singularly, whereas the same combination on the same allele is pathogenic in homozygous mode, or in heterozygous mode in association with another mutation; most likely, an heteroallelic combination of these two mutations is also pathogenic.
Although neither Thomsen2 nor Becker1 found that men are more frequently affected than women by MC, it has been suggested that myotonic symptoms may be more pronounced in men than in women.1 In our study, there was a clear disproportion in gender, with the 106 mutated MC probands comprising 73 male and 33 female subjects. A similar finding, albeit in a much smaller cohort, was recently reported.42
We did not detect mutations in 109 probands. One possible reason is that the appropriate clinical diagnosis had not been reached. Accordingly, we have started genetic screening for SCN4A gene, coding for the sodium channel voltage-gated type IV,17, 30 and found a mutation in 10 out of 15 probands investigated so far (data not shown). Another possibility could be related to the presence of macro-deletions and/or duplications and/or mutations in intronic or promoter regions of the CLCN1 gene, as these were not specifically investigated in this study with the methods used. Exon-wide deletion or duplication in the CLCN1 gene can now be performed by multiplex-ligation-dependent probe amplification, as recently reported by Raja Rayan et al.43
The number of novel additional mutations found in our study and their distribution throughout ClC-1 confirm the heterogeneity of MC pathogenesis. As also observed in other studies, most of the new mutations we detected appeared specific to individual patients or families. Nevertheless, our data suggest that, at least in the context of the Italian MC patient population, screening for CLCN1 mutations should initiate with exons 4 and 5 where a high mutation frequency apparently occurs. In addition, electrophysiological studies would be required to ascertain the actual effect of our new mutations on channel activity, improve the genotype–phenotype correlation and support drug design in the development of more specific ClC-1-targeted therapeutic approach.
Accession codes
References
Becker, P. Myotonia Congenita and Syndromes Associated with Myotonia, (Thieme: Stuttgart, 1977).
Thomsen, J. Tonische Krämpfe in willkürlich beweglichen Muskeln in Folge von ererbter psychischer disposition. Arch. Psychiatr. Nervenkr. 6, 702–718 (1876).
Lossin, C. & George, A. L. Jr. Myotonia congenita. Adv. Genet. 63, 25–55 (2008).
Lehmann-Horn, F., Jurkat-Rott, K. & Rüdel, R. Ulm Muscle Centre: diagnostics and therapy of muscle channelopathies—guidelines of the Ulm Muscle Centre. Acta Myol. 27, 98–113 (2008).
Lorenz, C., Meyer-Kleine, C., Steinmeyer, K., Koch, M. C. & Jentsch, T. J. Genomic organization of the human muscle chloride channel ClC-1 and analysis of novel mutations leading to Becker-type myotonia. Hum. Mol. Genet. 3, 941–946 (1994).
Dutzler, R., Campbell, E. B., Cadene, M., Chait, B. T. & MacKinnon, R. X-ray structure of a ClC chloride channel at 3.0 Å reveals the molecular basis of anion selectivity. Nature 415, 287–294 (2002).
Estévez, R., Pusch, M., Ferrer-Costa, C., Orozco, M. & Jentsch, T. J. Functional and structural conservation of CBS domains from CLC chloride channels. J. Physiol. 557, 363–378 (2004).
Kubisch, C., Schmidt-Rose, T., Fontaine, B., Bretag, A. H. & Jentsch, T. J. ClC-1 chloride channel mutations in myotonia congenita: variable penetrance of mutations shifting the voltage dependence. Hum. Mol. Genet. 7, 1753–1760 (1998).
Brugnoni, R., Galantini, S., Confalonieri, P., Balestrini, M. R., Cornelio, F. & Mantegazza, R. Identification of three novel mutations in the major human skeletal muscle chloride channel gene (CLCN1), causing myotonia congenita. Hum. Mutat. 14, 447 (1999).
de Diego, C., Gámez, J., Plassart-Schiess, E., Lasa, A., Del Río, E., Cervera, C. et al. Novel mutations in the muscle chloride channel CLCN1 gene causing myotonia congenita in Spanish families. J. Neurol. 246, 825–829 (1999).
Esteban, J., Neumeyer, A. M., McKenna-Yasek, D. & Brown, R. H. Identification of two mutations and a polymorphism in the chloride channel CLCN-1 in patients with Becker’s generalized myotonia. Neurogenetics 1, 185–188 (1998).
Fialho, D., Schorge, S., Pucovska, U., Davies, N. P., Labrum, R., Haworth, A. et al. Chloride channel myotonia: exon 8 hot-spot for dominant-negative interactions. Brain 130, 3265–3274 (2007).
Mazón, M. J., Barros, F., De la Peña, P., Quesada, J. F., Escudero, A., Cobo, A. M. et al. Screening for mutations in Spanish families with myotonia. Functional analysis of novel mutations in CLCN1 gene. Neuromuscul. Disord. 22, 231–243 (2012).
Modoni, A., D’Amico, A., Dallapiccola, B., Mereu, M. L., Merlini, L., Pagliarani, S. et al. Low-rate repetitive nerve stimulation protocol in an Italian cohort of patients affected by recessive myotonia congenita. J. Clin. Neurophysiol. 28, 39–44 (2011).
Pusch, M. Myotonia caused by mutations in the muscle chloride channel gene CLCN1. Hum. Mutat. 19, 423–434 (2002).
Sangiuolo, F., Botta, A., Mesoraca, A., Servidei, S., Merlini, L., Fratta, G. et al. Identification of five new mutations and three novel polymorphisms in the muscle chloride channel gene (CLCN1) in 20 Italian patients with dominant and recessive myotonia congenita. Hum. Mutat. 11, 331 (1998).
Trip, J., Drost, G., Verbove, D. J., van der Kooi, A. J., Kuks, J. B., Notermans, N. C. et al. In tandem analysis of CLCN1 and SCN4A greatly enhances mutation detection in families with non-dystrophic myotonia. Eur. J. Hum. Genet. 16, 921–929 (2008).
Wu, F. F., Ryan, A., Devaney, J., Warnstedt, M., Korade-Mirnics, Z., Poser, B. et al. Novel CLCN1 mutations with unique clinical and electrophysiological consequences. Brain 125, 2392–2407 (2002).
Leiden Open Variation Database. Chloride channel 1, skeletal muscle (CLCN1). http://chromium.liacs.nl/LOVD2/ (2012).
Plassart-Schiess, E., Gervais, A., Eymard, B., Lagueny, A., Pouget, J., Warter, J. M. et al. Novel muscle chloride channel (CLCN1) mutations in myotonia congenita with various modes of inheritance including incomplete dominance and penetrance. Neurology 50, 1176–1179 (1998).
Sloan Brown, K. & George, A. L. Inheritance of three distinct muscle chloride channel gene (CLCN1) mutations in a single recessive myotonia congenita family. Neurology 48, 542–543 (1997).
Sali, A., Potterton, L., Yuan, F., van Vlijmen, H. & Karplus, M. Evaluation of comparative protein modeling by MODELLER. Proteins 23, 318–326 (1995).
Hess, B., Kutzner, C., van der Spoel, D. & Lindahl, E. GROMACS 4: Algorithms for highly efficient, load-balanced, and scalable molecular simulation. J. Chem. Theory Comput. 4, 435–447 (2008).
Laskowski, R. A., MacArthur, M. W., Moss, D. S. & Thornton, J. M. PROCHECK—a program to check the stereochemical quality of protein structures. J. App. Cryst. 26, 283–291 (1993).
Zhang, C., Li, W. H., Krainer, A. R. & Zhang, M. Q. RNA landscape of evolution for optimal exon and intron discrimination. Proc. Natl. Acad. Sci. USA 105, 5797–5802 (2008).
Sironi, M., Menozzi, G., Riva, L., Cagliani, R., Comi, G. P., Bresolin, N. et al. Silencer elements as possible inhibitors of pseudoexon splicing. Nucleic Acid Res. 32, 1783–1791 (2004).
Shalata, A., Furman, H., Adir, V., Adir, N., Hujeirat, Y., Shalev, S. A. et al. Myotonia congenita in a large consanguineous Arab family: insight into the clinical spectrum of carriers and double heterozygotes of a novel mutation in the chloride channel CLCN1 gene. Muscle Nerve 41, 464–469 (2010).
Castro, P. M., Real, R., Leão, M. & Silveira, F. Co-dominant expression of a new mutation in myotonia congenita. Abstract. 21st Meeting of the European Neurological Society, (Lisbon, Portugal, 2011).
George, A. L. Jr, Sloan-Brown, K., Fenichel, G. M., Mitchell, G. A., Spiegel, R. & Pascuzzi, R. M. Nonsense and missense mutations of the muscle chloride channel gene in patients with myotonia congenita. Hum. Mol. Genet. 3, 2071–2072 (1994).
Lehmann-Horn, F., Mailander, V., Heine, R. & George, A. L. Myotonia levior is a chloride channel disorder. Hum. Mol. Genet. 4, 1397–1402 (1995).
Dupre, N., Chrestian, N., Bouchard, J. P., Rossignol, E., Brunet, D., Sternberg, D. et al. Clinical, electrophysiologic, and genetic study of non-dystrophic myotonia in French-Canadians. Neuromuscul. Disord. 19, 330–334 (2009).
Sun, C., Tranebjaerg, L., Torbergsen, T., Holmgren, G. & Van Ghelue, M. Spectrum of CLCN1 mutations in patients with myotonia congenita in Northern Scandinavia. Eur. J. Hum. Genet. 9, 903–909 (2001).
Fahlke, C. Molecular mechanisms of ion conduction in ClC-type chloride channels: lessons from disease-causing mutations. Kidney Int. 57, 780–786 (2000).
Meyer-Kleine, C., Steinmeyer, K., Ricker, K., Jentsch, T. J. & Koch, M. C. Spectrum of mutations in the major human skeletal muscle chloride channel gene (CLCN1) leading to myotonia. Am. J. Hum. Genet. 57, 1325–1334 (1995).
Zhang, J., Bendahhou, S., Sanguinetti, M. C. & Ptacek, L. J. Functional consequences of chloride channel gene (CLCN1) mutations causing myotonia congenita. Neurology 54, 937–942 (2000).
Fahlke, C., Desai, R. R., Gillani, N. & George, A. L. Jr. Residues lining the inner pore vestibule of human muscle chloride channels. J. Biol. Chem. 276, 1759–1765 (2001).
Desaphy, J.-F., Rolland, J.-F., Valente, E. M., LoMonaco, M. & Conte Camerino, D. Functional alteration of ClC-1 channel mutants associated with transient weakness in myotonia congenita. Biophys. J. 92, 273a (2007).
Hsiao, K. M., Huang, R. Y., Tang, P. H. & Lin, M. J. Functional study of CLC-1 mutants expressed in Xenopus oocytes reveals that a C-terminal region Thr891-Ser892-Thr893 is responsible for the effects of protein kinase C activator. Cell Physiol. Biochem. 25, 687–694 (2010).
Bennetts, B., Rychkov, G. Y., Ng, H. L., Morton, C. J., Stapleton, D., Parker, M. W. et al. Cytoplasmic ATP-sensing domains regulate gating of skeletal muscle ClC-1 chloride channels. J. Biol. Chem. 280, 32452–32458 (2005).
Macias, M. J., Teijido, O., Zifarelli, G., Martin, P., Ramirez-Espain, X., Zorzano, A. et al. Myotonia-related mutations in the distal C-terminus of ClC-1 and ClC-0 chloride channels affect the structure of a poly-proline helix. Biochem. J. 403, 79–87 (2007).
Ma, L., Rychkova, G. Y. & Bretag, A. H. Functional study of cytoplasmic loops of human skeletal muscle chloride channel, hClC-1. Int. J. Biochem. Cell Biol. 41, 1402–1409 (2009).
Tan, S. V., Matthews, E., Barber, M., Burge, J. A., Rajakulendran, S., Fialho, D. et al. Refined exercise testing can aid DNA-based diagnosis in muscle channelopathies. Ann. Neurol. 69, 328–340 (2011).
Raja Rayan, D. L., Haworth, A., Sud, R., Matthews, E., Fialho, D., Burge, J. et al. A new explanation for recessive myotonia congenita: Exon deletions and duplications in CLCN1. Neurology 78, 1953–1958 (2012).
Acknowledgements
We are grateful to the MC patients and their families for their cooperation, and to the doctors who provided the blood samples for the study. This work was supported by the Italian Ministry of Health (Grant No. 1580433).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on Journal of Human Genetics website
Supplementary information
Rights and permissions
About this article
Cite this article
Brugnoni, R., Kapetis, D., Imbrici, P. et al. A large cohort of myotonia congenita probands: novel mutations and a high-frequency mutation region in exons 4 and 5 of the CLCN1 gene. J Hum Genet 58, 581–587 (2013). https://doi.org/10.1038/jhg.2013.58
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/jhg.2013.58
