
Abstract
GroEL, a heat-shock protein that acts as a molecular chaperone1, is overproduced in endosymbiotic but not in free-living bacteria2,3,4, presumably to assist in the folding of conformationally damaged proteins. Here we show that the overproduction of GroEL in Escherichia coli masks the effects of harmful mutations that have accumulated during a simulated process of vertical transmission. This molecular mechanism, which may be an adaptation to the bacterium's intracellular lifestyle, is able to rescue lineages from a progressive fitness decline resulting from the fixation of deleterious mutations under strong genetic drift5,6.
Similar content being viewed by others

Main
Endosymbiotic bacteria have small population sizes, do not undergo recombination, and are maternally transmitted through tight population bottlenecks7, causing the fixation of deleterious mutations due to genetic drift and hence an irreversible decline in fitness8. However, endosymbiosis is surprisingly stable and persists over long periods9, which has led to the suggestion5 that groE (the GroEL-encoding operon) could be buffering the mutational loss of functionally active proteins because, unlike other endosymbiont genes, it is subject to strong purifying selection5.
To test this idea, we investigated the effects of overexpression of groE in a set of E. coli strains (a free-living bacterium close to several endosymbionts9) with mutations randomly accumulated throughout the genome. These spontaneous mutations were fixed by random genetic drift in a process that simulated the vertical transmission of a single endosymbiont between hosts.
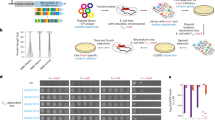
We studied the accumulation of mutations in 12 replicate lines of two E. coli B genotypes, one of which had a 3.3-fold-increased mutation rate10. These genotypes had already adapted to a simple environment (DM25 medium) for 10,000 generations11, suggesting that mutation accumulation might result in a decline in fitness. After 3,240 generations of mutation accumulation, we measured the fitness of the strains evolved on DM25 relative to their respective ancestors. As expected, the mutated strains lost fitness (Fig. 1a) as a result of the accumulation of deleterious mutations (W̄ = 0.5168 ± 0.0159 for mutators; W̄ =0.5208 ± 0.0087 for non-mutators; paired t-tests, P < 0.0001).

a, The fitness values for the ancestral strains of mutators and non-mutators are the same (left); 'mutated', fitness loss after 3,240 generations of mutation accumulation; 'groEc', fitness recovery of strains in which groE is overexpressed. Culture conditions, fitness assays and phenotypic markers are described elsewhere11, as is the mutation-accumulation protocol12. b, The regulated E. coli groE operon was changed to a constitutive form (groEc) by putting the structural genes under the control of the β-lactamase gene promoter (Pbla). Recombinant genotypes were obtained by transduction using P1vir (ref. 13). The groEc construct contains part of the flanking yjeH gene, the inducible tetracycline-resistance operon (tetA/R) for transductant selection, the Pbla promoter, and the groE coding regions (S and L). Restriction-enzyme cutting sites are indicated; P, promoter. Further details are available from the authors.
We then replaced the regulated groE operon of the mutated strains with a constitutive allele (groEc ; Fig. 1b). To estimate the fitness of the groEc strains, we ran competition experiments against the corresponding ancestors on DM25. Surprisingly, none of the groEc strains reached the expected cell density during growth overnight. A simple explanation could be that an overproduction of GroEL of about 86 ± 16-fold is deleterious because it diverts amino acids away from other cellular functions.
To test this, we grew each groEc strain and its ancestor in DM25 supplemented with increasing concentrations of tryptone (a mixture of peptides and amino acids). We found that groE overexpression was deleterious in the presence of small amounts of tryptone, but that there were no significant differences in cell density between groEc strains and their non-mutated ancestors at tryptone concentrations of 0.1% or higher. In endosymbionts, amino acids are not limiting because these are abundant in their intracellular environment9.
To determine whether fitness estimates depend on the environment, we estimated fitness on DM25 and on DM25 ± 0.1% tryptone. Both fitness estimates were correlated (partial correlation test, P < 0.0001).
Figure 1a shows that the average fitness of the groEc strains derived from non-mutator strains (W̄ = 0.8801 ± 0.0214) was 75.9% greater than that of the mutated strains (paired t-test, P < 0.0001), but was 12% less than that of the ancestors (P < 0.0002); the average fitness of the groEc strains derived from evolved mutators was W¯ =0.8152 ± 0.0167, which is 61.6% greater than that of the mutated strains (paired t-test, P < 0.0001) but 18.48% less than that of the ancestral strains (P < 0.0001).
Is fitness recovery a result of the buffering of deleterious effects by GroEL, or is it simply a general benefit associated with increased concentrations of GroEL? In favour of the first possibility, the advantage of GroEL overproduction is evident only in an amino-acid-rich environment; also, if mutation compensation is occurring, we would expect a positive correlation between the extents of fitness loss and recovery, as evidenced by their partial correlation (1-tailed test, P = 0.0089). We conclude that GroEL overexpression is likely to be of help in maintaining these endosymbionts by protecting them against the harmful effects of accumulated mutations.
References
Sigler, P. B. et al. Annu. Rev. Biochem. 67, 581–608 (1998).
Sato, S. & Ishikawa, H. J. Bacteriol. 179, 2300–2304 (1997).
Aksoy, S. Insect Mol. Biol. 4, 32–29 (1995).
Charles, H., Heddi, A., Guillaud, J., Nardon, C. & Nardon, P. Biochem. Biophys. Res. Commun. 239, 769–774 (1997).
Moran, N. A. Proc. Natl Acad. Sci. USA 93, 2873–2878 (1996).
Rutherford, S. L. & Lindquist, S. Nature 396, 336–342 (1998).
Funk, D. J., Wernegreen, J. J. & Moran, N. A. Genetics 157, 477–489 (2001).
Muller, H. J. Mutat. Res. 1, 2–9 (1964).
Moran, N. A. & Wernegreen, J. J. Trends Ecol. Evol. 15, 321–326 (2000).
de Visser, J. A. G. M., Zeyl, C., Gerrish, P. J., Blanchard, J. L. & Lenski, R. E. Science 283, 404–406 (1999).
Lenski, R. E., Rose, M. R., Simpson, S. C. & Tadler, S. C. Am. Nat. 138, 1315–1341 (1991).
Kibota, T. T. & Lynch, M. Nature 381, 694–696 (1996).
Zyskind, J. W. & Bernstein, S. I. Recombinant DNA Laboratory Manual (Academic, San Diego, 1989).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
About this article
Cite this article
Fares, M., Ruiz-González, M., Moya, A. et al. GroEL buffers against deleterious mutations. Nature 417, 398 (2002). https://doi.org/10.1038/417398a
Issue Date:
DOI: https://doi.org/10.1038/417398a
