

A novel class of evolutionarily conserved genes defined by piwi are essential for stem cell self-renewal
- PMID: 9851978
- PMCID: PMC317255
- DOI: 10.1101/gad.12.23.3715
A novel class of evolutionarily conserved genes defined by piwi are essential for stem cell self-renewal
Abstract
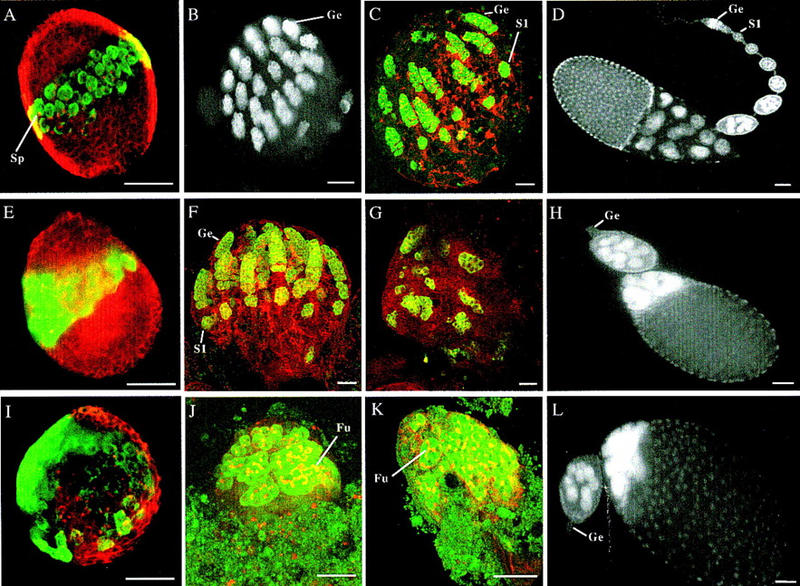
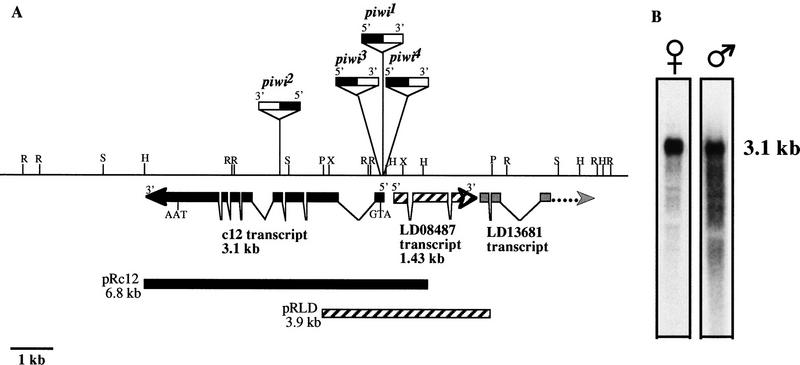
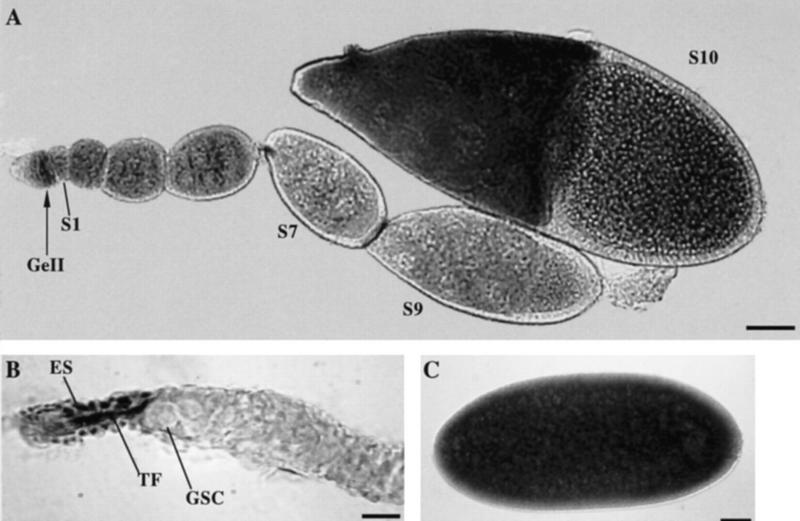
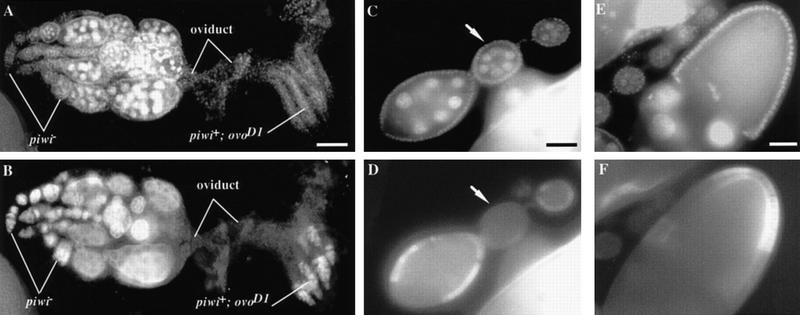
Germ-line stem cells (GSCs) serve as the source for gametogenesis in diverse organisms. We cloned and characterized the Drosophila piwi gene and showed that it is required for the asymmetric division of GSCs to produce and maintain a daughter GSC but is not essential for the further differentiation of the committed daughter cell. Genetic mosaic and RNA in situ analyses suggest that piwi expression in adjacent somatic cells regulates GSC division. piwi encodes a highly basic novel protein, well conserved during evolution. We isolated piwi homologs in Caenorhabditis elegans and humans and also identified Arabidopsis piwi-like genes known to be required for meristem cell maintenance. Decreasing C. elegans piwi expression reduces the proliferation of GSC-equivalent cells. Thus, piwi represents a novel class of genes required for GSC division in diverse organisms.
Figures


Similar articles
-
A Drosophila chromatin factor interacts with the Piwi-interacting RNA mechanism in niche cells to regulate germline stem cell self-renewal.Genetics. 2010 Oct;186(2):573-83. doi: 10.1534/genetics.110.119081. Epub 2010 Jul 20. Genetics. 2010. PMID: 20647505 Free PMC article.
-
Regulatory relationship among piwi, pumilio, and bag-of-marbles in Drosophila germline stem cell self-renewal and differentiation.Curr Biol. 2005 Jan 26;15(2):171-8. doi: 10.1016/j.cub.2005.01.005. Curr Biol. 2005. PMID: 15668175
-
piwi encodes a nucleoplasmic factor whose activity modulates the number and division rate of germline stem cells.Development. 2000 Feb;127(3):503-14. doi: 10.1242/dev.127.3.503. Development. 2000. PMID: 10631171
-
Uniting germline and stem cells: the function of Piwi proteins and the piRNA pathway in diverse organisms.Annu Rev Genet. 2011;45:447-69. doi: 10.1146/annurev-genet-110410-132541. Epub 2011 Sep 19. Annu Rev Genet. 2011. PMID: 21942366 Free PMC article. Review.
-
The tao of stem cells in the germline.Annu Rev Genet. 1997;31:455-91. doi: 10.1146/annurev.genet.31.1.455. Annu Rev Genet. 1997. PMID: 9442904 Review.
Cited by
-
Extremely stable Piwi-induced gene silencing in Caenorhabditis elegans.EMBO J. 2012 Aug 15;31(16):3422-30. doi: 10.1038/emboj.2012.213. Epub 2012 Jul 31. EMBO J. 2012. PMID: 22850670 Free PMC article.
-
Profiling of the small RNA populations in human testicular germ cell tumors shows global loss of piRNAs.Mol Cancer. 2015 Aug 12;14:153. doi: 10.1186/s12943-015-0411-4. Mol Cancer. 2015. PMID: 26265322 Free PMC article.
-
Promoters recognized by forkhead proteins exist for individual 21U-RNAs.Mol Cell. 2012 Sep 14;47(5):734-45. doi: 10.1016/j.molcel.2012.06.021. Epub 2012 Jul 19. Mol Cell. 2012. PMID: 22819322 Free PMC article.
-
Zili inhibits transforming growth factor-beta signaling by interacting with Smad4.J Biol Chem. 2010 Feb 5;285(6):4243-4250. doi: 10.1074/jbc.M109.079533. Epub 2009 Dec 9. J Biol Chem. 2010. PMID: 20007318 Free PMC article.
-
Germ Cell Lineage Homeostasis in Drosophila Requires the Vasa RNA Helicase.Genetics. 2019 Nov;213(3):911-922. doi: 10.1534/genetics.119.302558. Epub 2019 Sep 4. Genetics. 2019. PMID: 31484689 Free PMC article.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Brown EH, King RC. Oogonial and spermatogonial differentiation within a mosaic gonad of Drosophila melanogaster. Growth. 1962;26:53–70. - PubMed
-
- ————— Studies on the events resulting in the formation of an egg chamber in Drosophila melanogaster. Growth. 1964;28:41–81. - PubMed
-
- Brown NH, Kafatos FC. Functional libraries from Drosophila embryos. J Mol Biol. 1988;203:425–437. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials