

HIV-1 directly kills CD4+ T cells by a Fas-independent mechanism
- PMID: 9529327
- PMCID: PMC2212217
- DOI: 10.1084/jem.187.7.1113
HIV-1 directly kills CD4+ T cells by a Fas-independent mechanism
Abstract
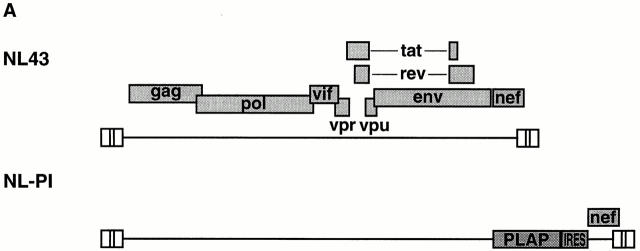
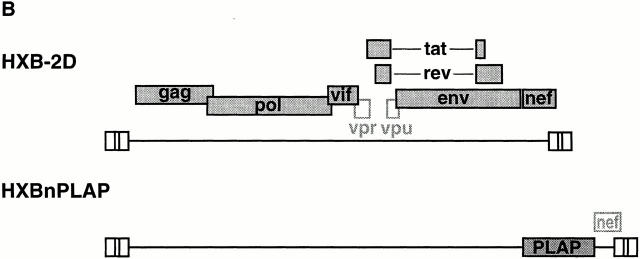
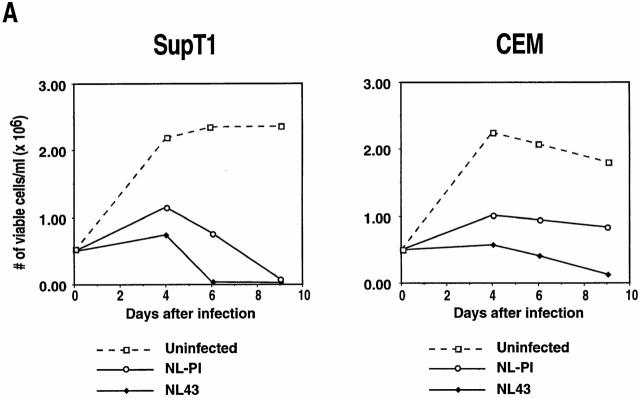
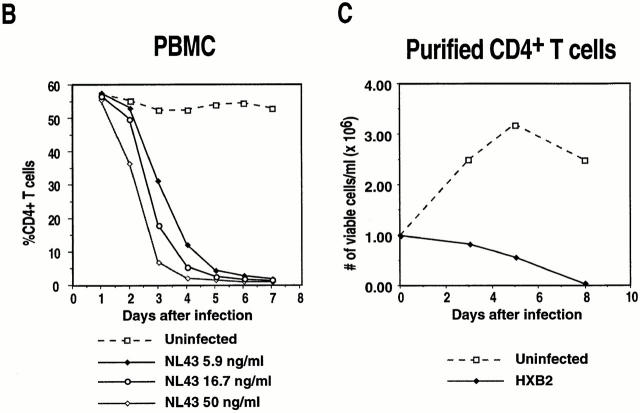
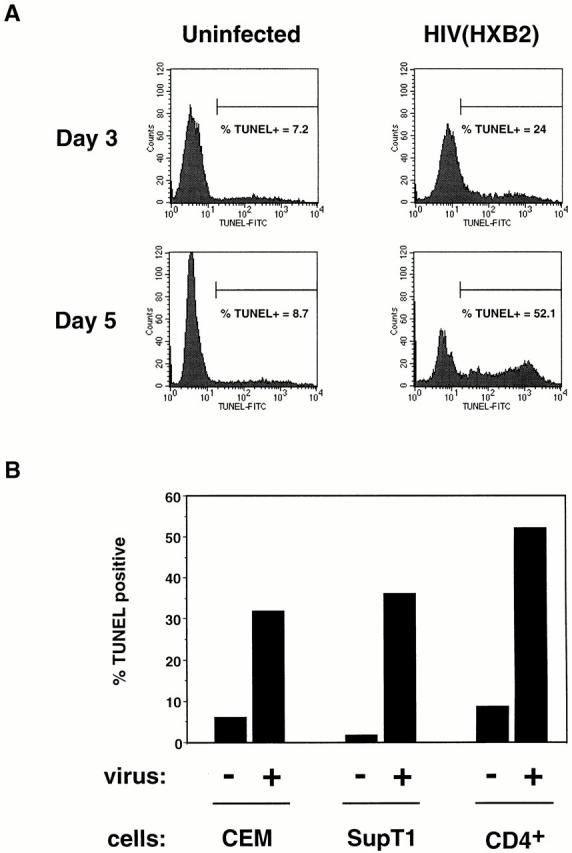
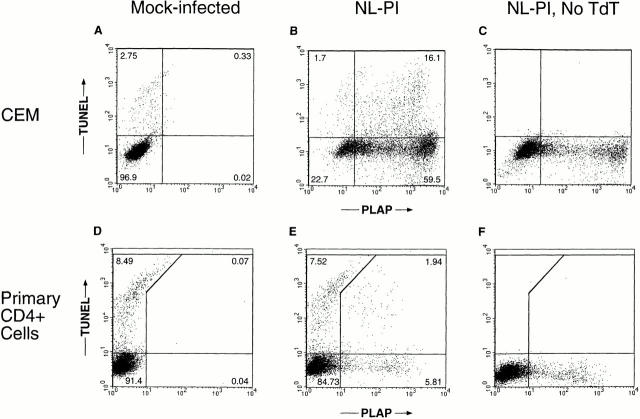
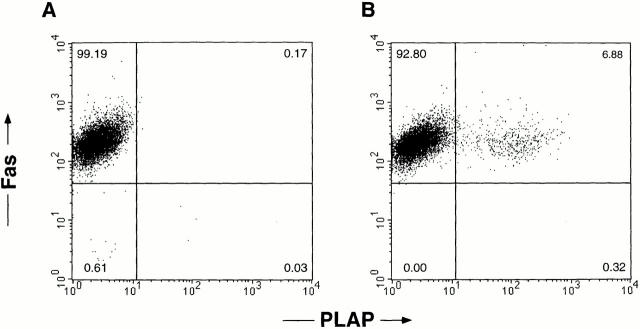
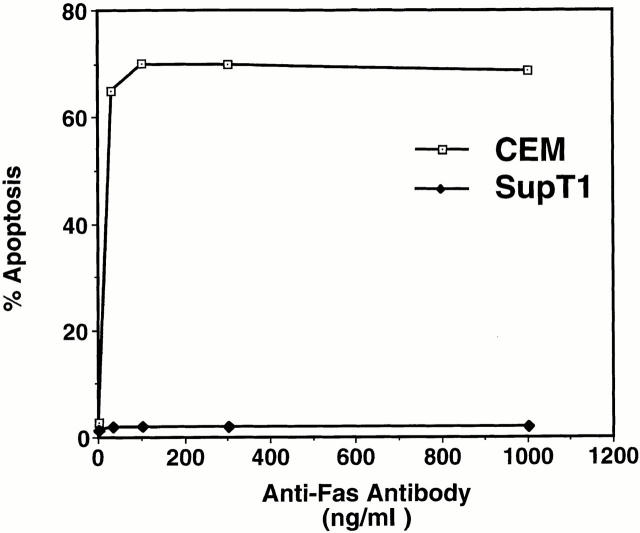
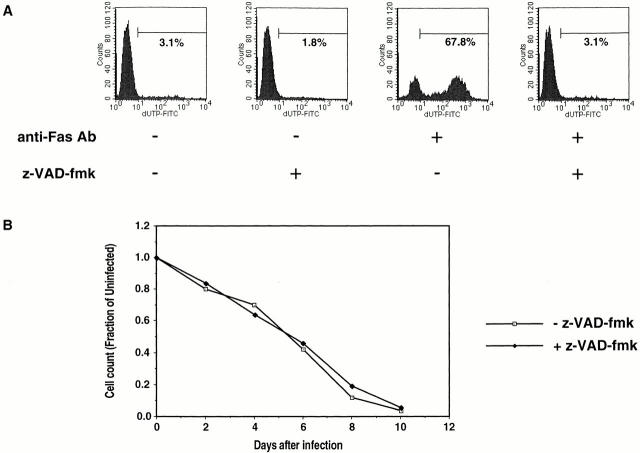
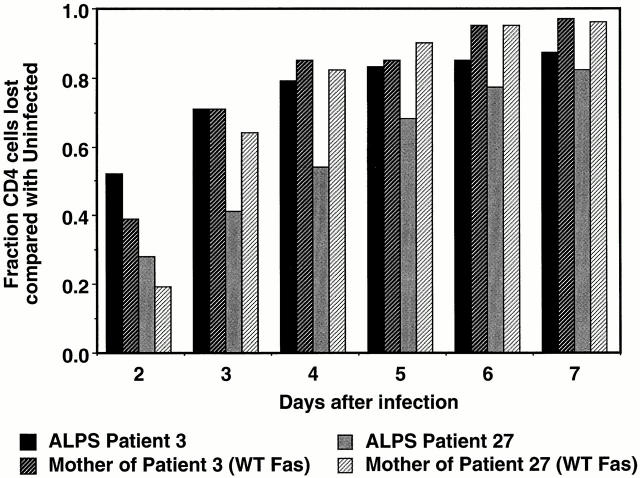
The mechanism by which HIV-1 induces CD4(+) T cell death is not known. A fundamental issue is whether HIV-1 primarily induces direct killing of infected cells or indirectly causes death of uninfected bystander cells. This question was studied using a reporter virus system in which infected cells are marked with the cell surface protein placental alkaline phosphatase (PLAP). Infection by HIV-PLAP of peripheral blood mononuclear cells (PBMCs) and T cell lines leads to rapid depletion of CD4(+) T cells and induction of apoptosis. The great majority of HIV-induced T cell death in vitro involves direct loss of infected cells rather than indirect effects on uninfected bystander cells. Because of its proposed role in HIV-induced cell death, we also examined the Fas (CD95/Apo1) pathway in killing of T cells by HIV-1. Infected PBMCs or CEM cells display no increase in surface Fas relative to uninfected cells. In addition, HIV-1 kills CEM and Jurkat T cells in the presence of a caspase inhibitor that completely blocks Fas-mediated apoptosis. HIV-1 also depletes CD4+ T cells in PBMCs from patients who have a genetically defective Fas pathway. These results suggest that HIV-1 induces direct apoptosis of infected cells and kills T cells by a Fas-independent mechanism.
Figures
Similar articles
-
CCR5 mediates Fas- and caspase-8 dependent apoptosis of both uninfected and HIV infected primary human CD4 T cells.AIDS. 2002 Jul 26;16(11):1467-78. doi: 10.1097/00002030-200207260-00003. AIDS. 2002. PMID: 12131184
-
Productive HIV-1 infection of primary CD4+ T cells induces mitochondrial membrane permeabilization leading to a caspase-independent cell death.J Biol Chem. 2002 Jan 11;277(2):1477-87. doi: 10.1074/jbc.M102671200. Epub 2001 Oct 31. J Biol Chem. 2002. PMID: 11689551
-
Type I interferon upregulates Bak and contributes to T cell loss during human immunodeficiency virus (HIV) infection.PLoS Pathog. 2013;9(10):e1003658. doi: 10.1371/journal.ppat.1003658. Epub 2013 Oct 10. PLoS Pathog. 2013. PMID: 24130482 Free PMC article. Clinical Trial.
-
Role of FAS in HIV infection.Curr HIV Res. 2003 Oct;1(4):405-17. doi: 10.2174/1570162033485131. Curr HIV Res. 2003. PMID: 15049427 Review.
-
Mechanisms of CD4+ T lymphocyte cell death in human immunodeficiency virus infection and AIDS.J Gen Virol. 2003 Jul;84(Pt 7):1649-1661. doi: 10.1099/vir.0.19110-0. J Gen Virol. 2003. PMID: 12810858 Review.
Cited by
-
Death of CD4(+) T-cell lines caused by human immunodeficiency virus type 1 does not depend on caspases or apoptosis.J Virol. 2002 May;76(10):5094-107. doi: 10.1128/jvi.76.10.5094-5107.2002. J Virol. 2002. PMID: 11967325 Free PMC article.
-
Wnt/β-Catenin Protects Lymphocytes from HIV-Mediated Apoptosis via Induction of Bcl-xL.Viruses. 2022 Jul 2;14(7):1469. doi: 10.3390/v14071469. Viruses. 2022. PMID: 35891449 Free PMC article.
-
Early reduction of the over-expression of CD40L, OX40 and Fas on T cells in HIV-1 infection during triple anti-retroviral therapy: possible implications for lymphocyte traffic and functional recovery.Clin Exp Immunol. 1999 May;116(2):307-15. doi: 10.1046/j.1365-2249.1999.00872.x. Clin Exp Immunol. 1999. PMID: 10337023 Free PMC article.
-
Making a Monkey out of Human Immunodeficiency Virus/Simian Immunodeficiency Virus Pathogenesis: Immune Cell Depletion Experiments as a Tool to Understand the Immune Correlates of Protection and Pathogenicity in HIV Infection.Viruses. 2024 Jun 17;16(6):972. doi: 10.3390/v16060972. Viruses. 2024. PMID: 38932264 Free PMC article. Review.
-
The Hitchhiker Guide to CD4+ T-Cell Depletion in Lentiviral Infection. A Critical Review of the Dynamics of the CD4+ T Cells in SIV and HIV Infection.Front Immunol. 2021 Jul 21;12:695674. doi: 10.3389/fimmu.2021.695674. eCollection 2021. Front Immunol. 2021. PMID: 34367156 Free PMC article.
References
-
- Ho DD, Neumann AU, Perelson AS, Chen W, Leonard JM, Markowitz M. Rapid turnover of plasma virions and lymphocytes in HIV-1 infection. Nature. 1995;373:123–126. - PubMed
-
- Wei X, Ghosh SK, Taylor ME, Johnson VA, Emini EA, Deutsch P, Lifson JD, Bonhoeffer S, Nowak MA, Hahn BH, et al. Viral dynamics in human immunodeficiency virus type 1 infection. Nature. 1995;373:117–122. - PubMed
-
- Perelson AS, Neumann AU, Markowitz M, Leonard JM, Ho DD. HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science. 1996;271:1582–1586. - PubMed
-
- Mellors JW, Rinaldo C, Gupta P, White RM, Todd JA, Kingsley LA. Prognosis in HIV-1 infection predicted by the quantity of virus in plasma. Science. 1996;272:1167–1170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
