

Bone marrow adipogenic lineage precursors promote osteoclastogenesis in bone remodeling and pathologic bone loss
- PMID: 33206630
- PMCID: PMC7810488
- DOI: 10.1172/JCI140214
Bone marrow adipogenic lineage precursors promote osteoclastogenesis in bone remodeling and pathologic bone loss
Abstract
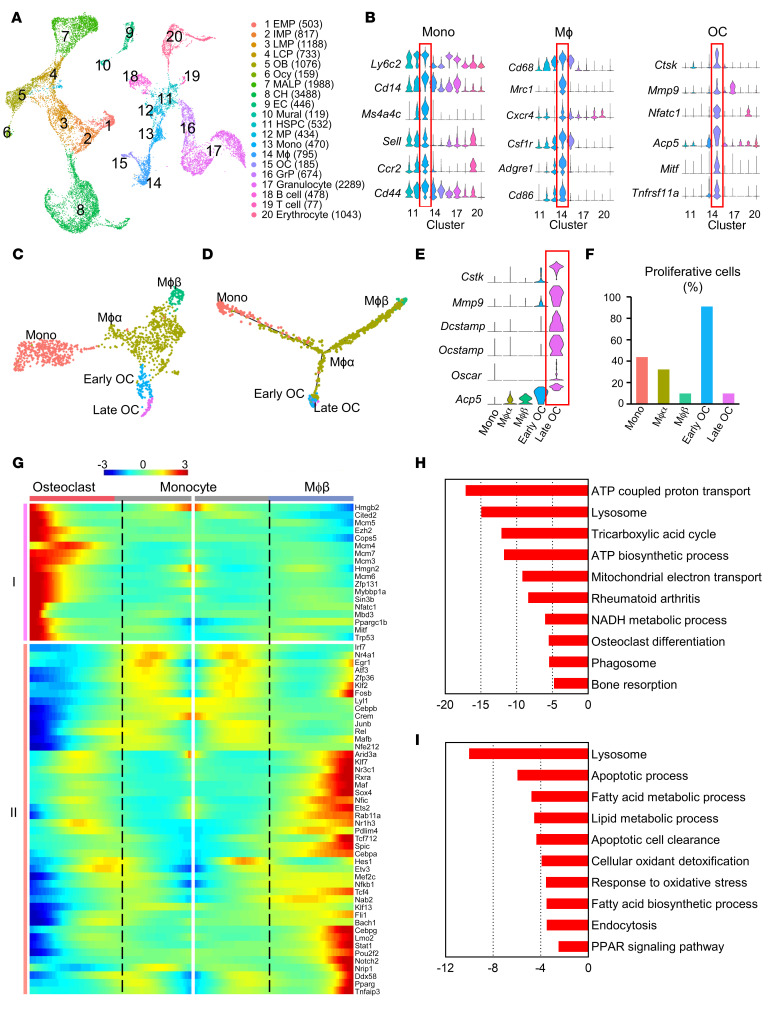
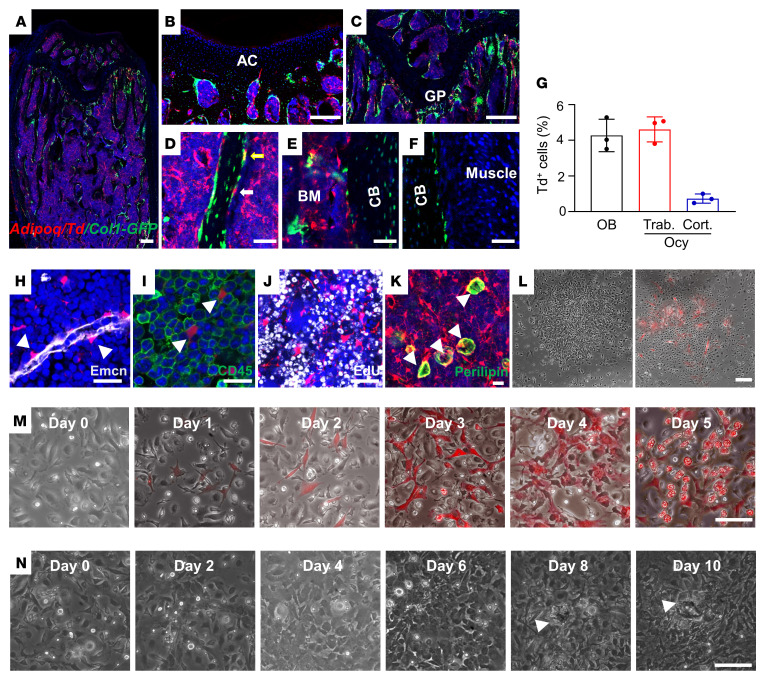
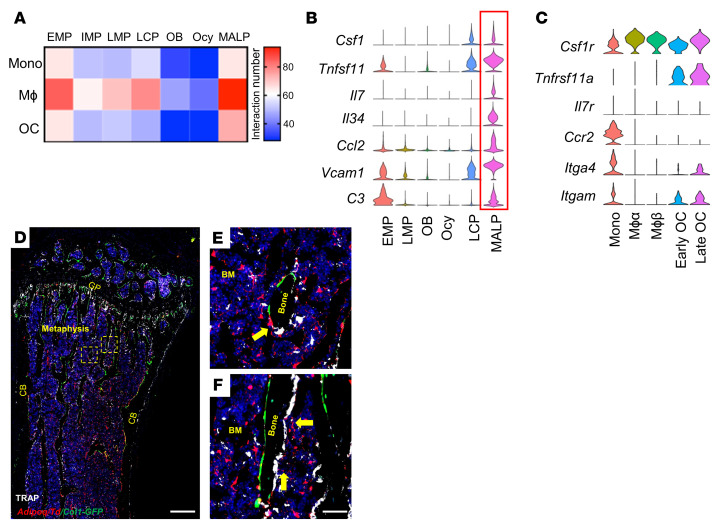
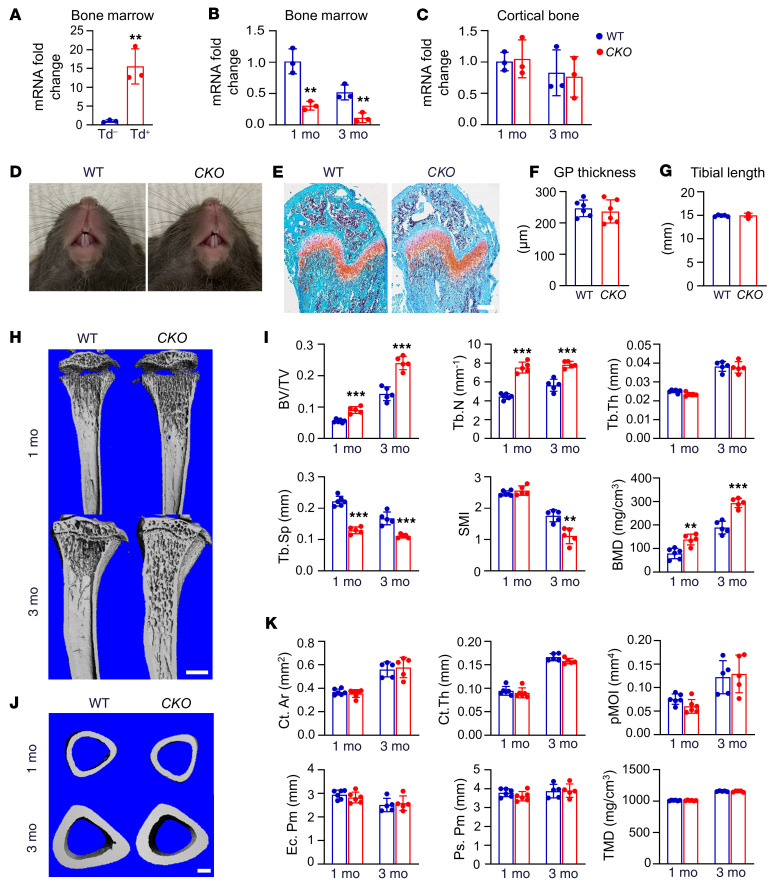
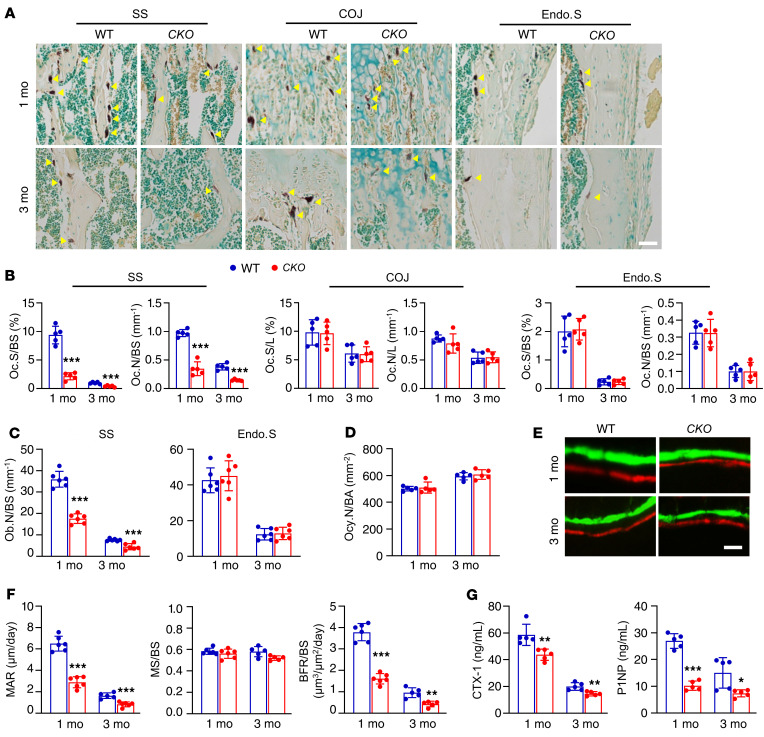
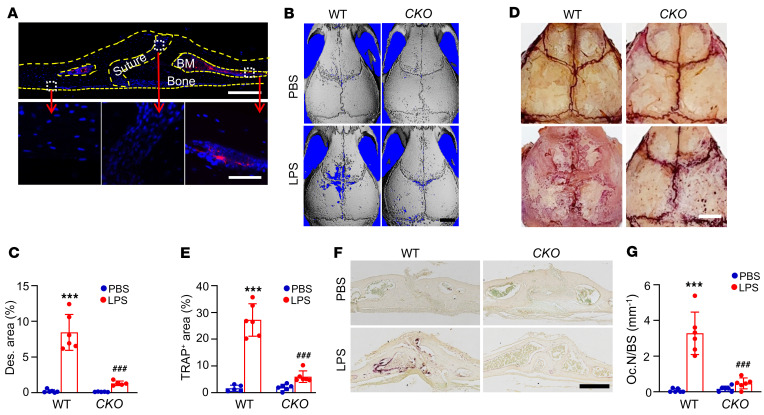
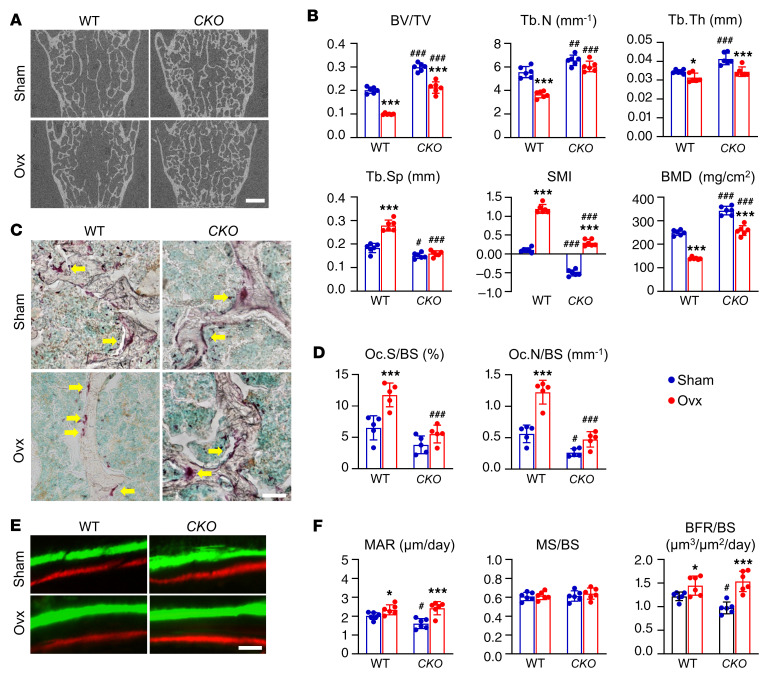
Bone is maintained by coupled activities of bone-forming osteoblasts/osteocytes and bone-resorbing osteoclasts. Alterations in this relationship can lead to pathologic bone loss such as osteoporosis. It is well known that osteogenic cells support osteoclastogenesis via production of RANKL. Interestingly, our recently identified bone marrow mesenchymal cell population-marrow adipogenic lineage precursors (MALPs) that form a multidimensional cell network in bone-was computationally demonstrated to be the most interactive with monocyte-macrophage lineage cells through high and specific expression of several osteoclast regulatory factors, including RANKL. Using an adipocyte-specific Adipoq-Cre to label MALPs, we demonstrated that mice with RANKL deficiency in MALPs have a drastic increase in trabecular bone mass in long bones and vertebrae starting from 1 month of age, while their cortical bone appears normal. This phenotype was accompanied by diminished osteoclast number and attenuated bone formation at the trabecular bone surface. Reduced RANKL signaling in calvarial MALPs abolished osteolytic lesions after LPS injections. Furthermore, in ovariectomized mice, elevated bone resorption was partially attenuated by RANKL deficiency in MALPs. In summary, our studies identified MALPs as a critical player in controlling bone remodeling during normal bone metabolism and pathological bone loss in a RANKL-dependent fashion.
Keywords: Bone Biology; Bone marrow differentiation.
Conflict of interest statement
Figures
Comment in
-
A critical relationship between bone and fat: the role of bone marrow adipose-derived RANKL in bone metabolism.EMBO Rep. 2021 Jul 5;22(7):e52986. doi: 10.15252/embr.202152986. Epub 2021 Jun 13. EMBO Rep. 2021. PMID: 34121307 Free PMC article.
Similar articles
-
Bone marrow adipogenic lineage precursors are the major regulator of bone resorption in adult mice.Res Sq [Preprint]. 2024 Aug 28:rs.3.rs-4809633. doi: 10.21203/rs.3.rs-4809633/v1. Res Sq. 2024. PMID: 39257979 Free PMC article. Preprint.
-
Transglutaminases factor XIII-A and TG2 regulate resorption, adipogenesis and plasma fibronectin homeostasis in bone and bone marrow.Cell Death Differ. 2017 May;24(5):844-854. doi: 10.1038/cdd.2017.21. Epub 2017 Apr 7. Cell Death Differ. 2017. PMID: 28387755 Free PMC article.
-
Csf1 from marrow adipogenic precursors is required for osteoclast formation and hematopoiesis in bone.Elife. 2023 Feb 13;12:e82112. doi: 10.7554/eLife.82112. Elife. 2023. PMID: 36779854 Free PMC article.
-
Control of RANKL gene expression.Bone. 2010 Apr;46(4):911-9. doi: 10.1016/j.bone.2009.08.050. Epub 2009 Aug 27. Bone. 2010. PMID: 19716455 Free PMC article. Review.
-
Osteoclast differentiation by RANKL and OPG signaling pathways.J Bone Miner Metab. 2021 Jan;39(1):19-26. doi: 10.1007/s00774-020-01162-6. Epub 2020 Oct 20. J Bone Miner Metab. 2021. PMID: 33079279 Review.
Cited by
-
Pericytes as the Orchestrators of Vasculature and Adipogenesis.Genes (Basel). 2024 Jan 19;15(1):126. doi: 10.3390/genes15010126. Genes (Basel). 2024. PMID: 38275607 Free PMC article. Review.
-
IFT20 governs mesenchymal stem cell fate through positively regulating TGF-β-Smad2/3-Glut1 signaling mediated glucose metabolism.Redox Biol. 2022 Aug;54:102373. doi: 10.1016/j.redox.2022.102373. Epub 2022 Jun 20. Redox Biol. 2022. PMID: 35751983 Free PMC article.
-
Flavonoid Phloretin Inhibits Adipogenesis and Increases OPG Expression in Adipocytes Derived from Human Bone-Marrow Mesenchymal Stromal-Cells.Nutrients. 2021 Nov 22;13(11):4185. doi: 10.3390/nu13114185. Nutrients. 2021. PMID: 34836440 Free PMC article.
-
Skeletal Stem Cells as the Developmental Origin of Cellular Niches for Hematopoietic Stem and Progenitor Cells.Curr Top Microbiol Immunol. 2021;434:1-31. doi: 10.1007/978-3-030-86016-5_1. Curr Top Microbiol Immunol. 2021. PMID: 34850280 Free PMC article.
-
A systematic dissection of human primary osteoblasts in vivo at single-cell resolution.Aging (Albany NY). 2021 Aug 24;13(16):20629-20650. doi: 10.18632/aging.203452. Epub 2021 Aug 24. Aging (Albany NY). 2021. PMID: 34428745 Free PMC article.
References
-
- Kains, JA on behalf of the World Health Organisation Scientific Group. Assessment of osteoporosis at the primary health care level. WHO Collaborating Centre for Metabolic Bone Diseases, University of Sheffield 2007. https://www.sheffield.ac.uk/FRAX/pdfs/WHO_Technical_Report.pdf.
-
- Kim N, Odgren PR, Kim DK, Marks SC, Choi Y. Diverse roles of the tumor necrosis factor family member TRANCE in skeletal physiology revealed by TRANCE deficiency and partial rescue by a lymphocyte-expressed TRANCE transgene. Proc Natl Acad Sci U S A. 2000;97(20):10905–10910. doi: 10.1073/pnas.200294797. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
