

This is a preprint.
Furin Cleavage Site Is Key to SARS-CoV-2 Pathogenesis
- PMID: 32869021
- PMCID: PMC7457603
- DOI: 10.1101/2020.08.26.268854
Furin Cleavage Site Is Key to SARS-CoV-2 Pathogenesis
Update in
-
Loss of furin cleavage site attenuates SARS-CoV-2 pathogenesis.Nature. 2021 Mar;591(7849):293-299. doi: 10.1038/s41586-021-03237-4. Epub 2021 Jan 25. Nature. 2021. PMID: 33494095 Free PMC article.
Abstract
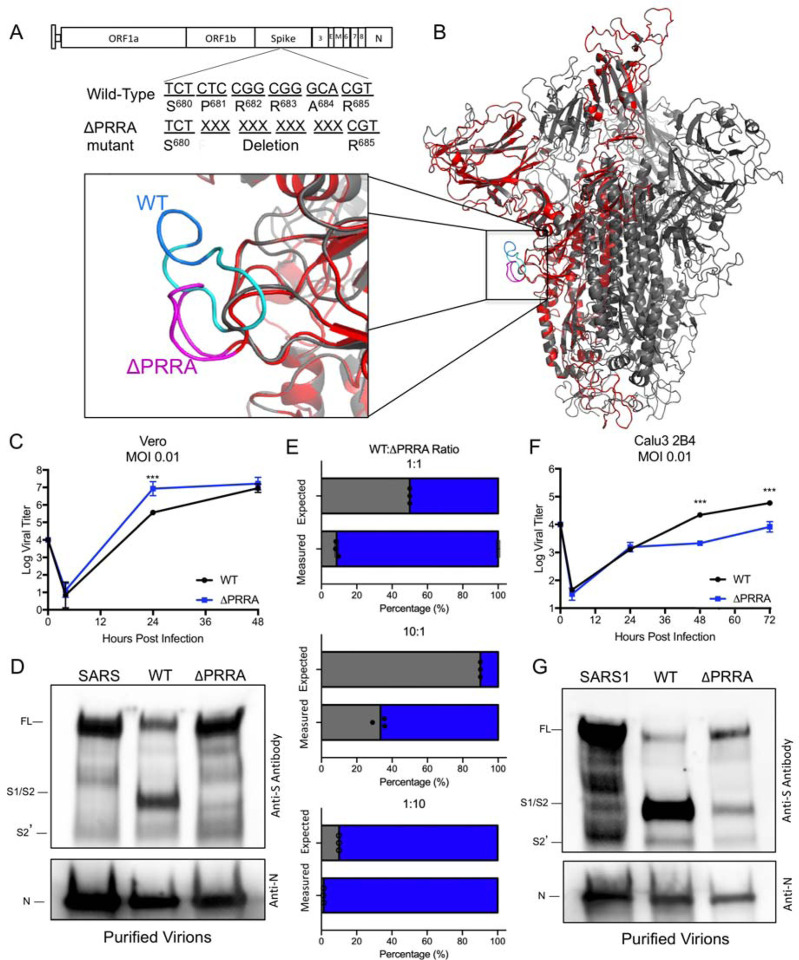
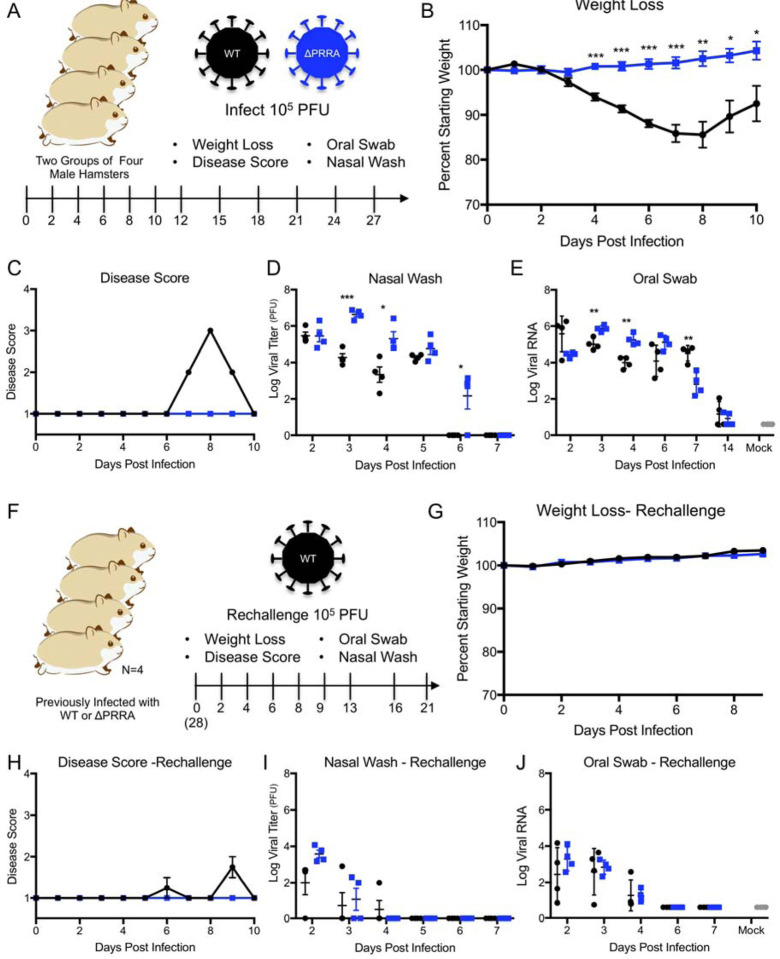
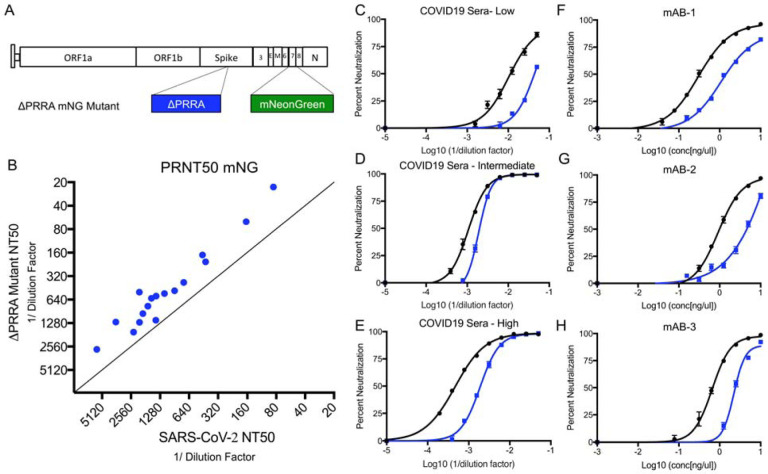
SARS-CoV-2 has resulted in a global pandemic and shutdown economies around the world. Sequence analysis indicates that the novel coronavirus (CoV) has an insertion of a furin cleavage site (PRRAR) in its spike protein. Absent in other group 2B CoVs, the insertion may be a key factor in the replication and virulence of SARS-CoV-2. To explore this question, we generated a SARS-CoV-2 mutant lacking the furin cleavage site (ΔPRRA) in the spike protein. This mutant virus replicated with faster kinetics and improved fitness in Vero E6 cells. The mutant virus also had reduced spike protein processing as compared to wild-type SARS-CoV-2. In contrast, the ΔPRRA had reduced replication in Calu3 cells, a human respiratory cell line, and had attenuated disease in a hamster pathogenesis model. Despite the reduced disease, the ΔPRRA mutant offered robust protection from SARS-CoV-2 rechallenge. Importantly, plaque reduction neutralization tests (PRNT 50 ) with COVID-19 patient sera and monoclonal antibodies against the receptor-binding domain found a shift, with the mutant virus resulting in consistently reduced PRNT 50 titers. Together, these results demonstrate a critical role for the furin cleavage site insertion in SARS-CoV-2 replication and pathogenesis. In addition, these findings illustrate the importance of this insertion in evaluating neutralization and other downstream SARS-CoV-2 assays.
Importance: As COVID-19 has impacted the world, understanding how SARS-CoV-2 replicates and causes virulence offers potential pathways to disrupt its disease. By removing the furin cleavage site, we demonstrate the importance of this insertion to SARS-CoV-2 replication and pathogenesis. In addition, the findings with Vero cells indicate the likelihood of cell culture adaptations in virus stocks that can influence reagent generation and interpretation of a wide range of data including neutralization and drug efficacy. Overall, our work highlights the importance of this key motif in SARS-CoV-2 infection and pathogenesis.
Article summary: A deletion of the furin cleavage site in SARS-CoV-2 amplifies replication in Vero cells, but attenuates replication in respiratory cells and pathogenesis in vivo. Loss of the furin site also reduces susceptibility to neutralization in vitro .
Conflict of interest statement
Competing interests
X.X., V.D.M., and P.-Y.S. have filed a patent on the reverse genetic system and reporter SARS-CoV-2. Other authors declare no competing interests.
Figures
Similar articles
-
Loss of furin cleavage site attenuates SARS-CoV-2 pathogenesis.Nature. 2021 Mar;591(7849):293-299. doi: 10.1038/s41586-021-03237-4. Epub 2021 Jan 25. Nature. 2021. PMID: 33494095 Free PMC article.
-
Identification of Common Deletions in the Spike Protein of Severe Acute Respiratory Syndrome Coronavirus 2.J Virol. 2020 Aug 17;94(17):e00790-20. doi: 10.1128/JVI.00790-20. Print 2020 Aug 17. J Virol. 2020. PMID: 32571797 Free PMC article.
-
QTQTN motif upstream of the furin-cleavage site plays a key role in SARS-CoV-2 infection and pathogenesis.Proc Natl Acad Sci U S A. 2022 Aug 9;119(32):e2205690119. doi: 10.1073/pnas.2205690119. Epub 2022 Jul 26. Proc Natl Acad Sci U S A. 2022. PMID: 35881779 Free PMC article.
-
Roles of the polybasic furin cleavage site of spike protein in SARS-CoV-2 replication, pathogenesis, and host immune responses and vaccination.J Med Virol. 2022 May;94(5):1815-1820. doi: 10.1002/jmv.27539. Epub 2021 Dec 31. J Med Virol. 2022. PMID: 34936124 Review.
-
SARS-CoV replication and pathogenesis in an in vitro model of the human conducting airway epithelium.Virus Res. 2008 Apr;133(1):33-44. doi: 10.1016/j.virusres.2007.03.013. Epub 2007 Apr 23. Virus Res. 2008. PMID: 17451829 Free PMC article. Review.
Cited by
-
The Biological Functions and Clinical Significance of SARS-CoV-2 Variants of Corcern.Front Med (Lausanne). 2022 May 20;9:849217. doi: 10.3389/fmed.2022.849217. eCollection 2022. Front Med (Lausanne). 2022. PMID: 35669924 Free PMC article. Review.
-
First Detection of SARS-CoV-2 B.1.617.2 (Delta) Variant of Concern in a Symptomatic Cat in Spain.Front Vet Sci. 2022 Apr 1;9:841430. doi: 10.3389/fvets.2022.841430. eCollection 2022. Front Vet Sci. 2022. PMID: 35433922 Free PMC article.
-
The Variation of SARS-CoV-2 and Advanced Research on Current Vaccines.Front Med (Lausanne). 2022 Jan 18;8:806641. doi: 10.3389/fmed.2021.806641. eCollection 2021. Front Med (Lausanne). 2022. PMID: 35118097 Free PMC article. Review.
-
SARS-CoV-2 tropism, entry, replication, and propagation: Considerations for drug discovery and development.PLoS Pathog. 2021 Feb 17;17(2):e1009225. doi: 10.1371/journal.ppat.1009225. eCollection 2021 Feb. PLoS Pathog. 2021. PMID: 33596266 Free PMC article. Review.
-
PTSD as an Endothelial Disease: Insights From COVID-19.Front Cell Neurosci. 2021 Oct 29;15:770387. doi: 10.3389/fncel.2021.770387. eCollection 2021. Front Cell Neurosci. 2021. PMID: 34776871 Free PMC article. Review.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous