

Developmental origin, functional maintenance and genetic rescue of osteoclasts
- PMID: 30971820
- PMCID: PMC6910203
- DOI: 10.1038/s41586-019-1105-7
Developmental origin, functional maintenance and genetic rescue of osteoclasts
Abstract
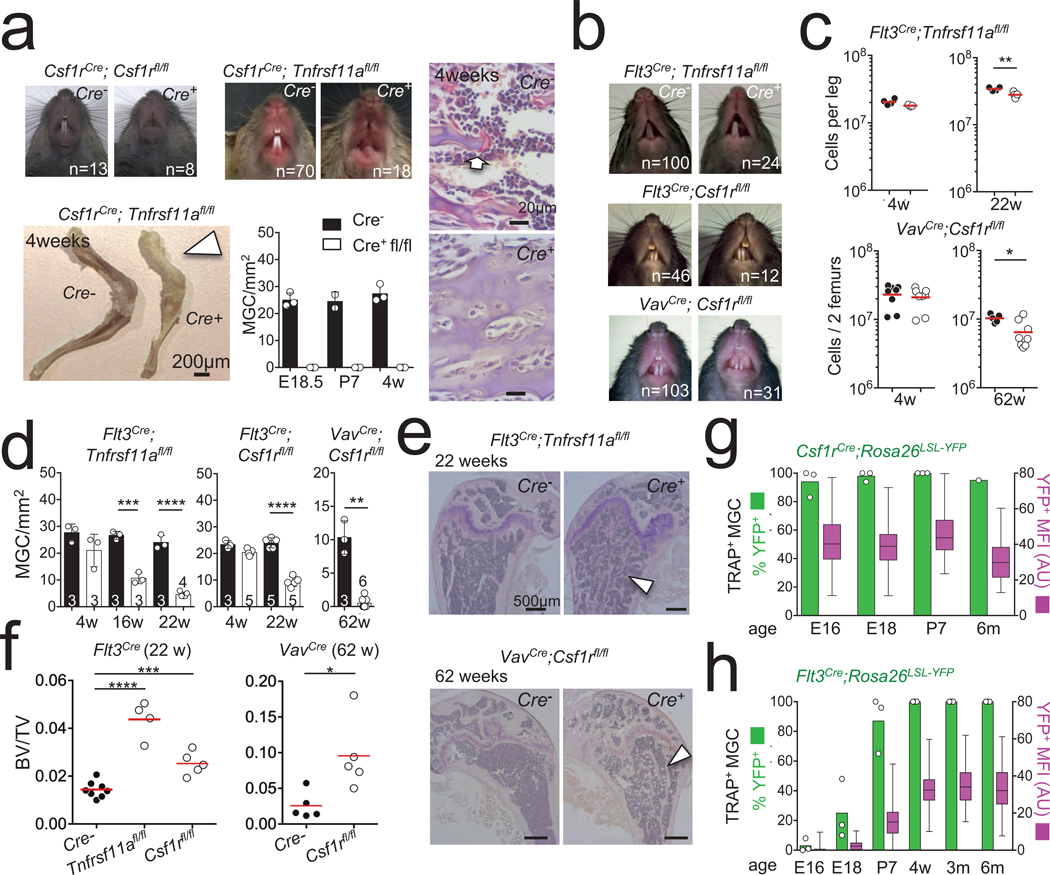
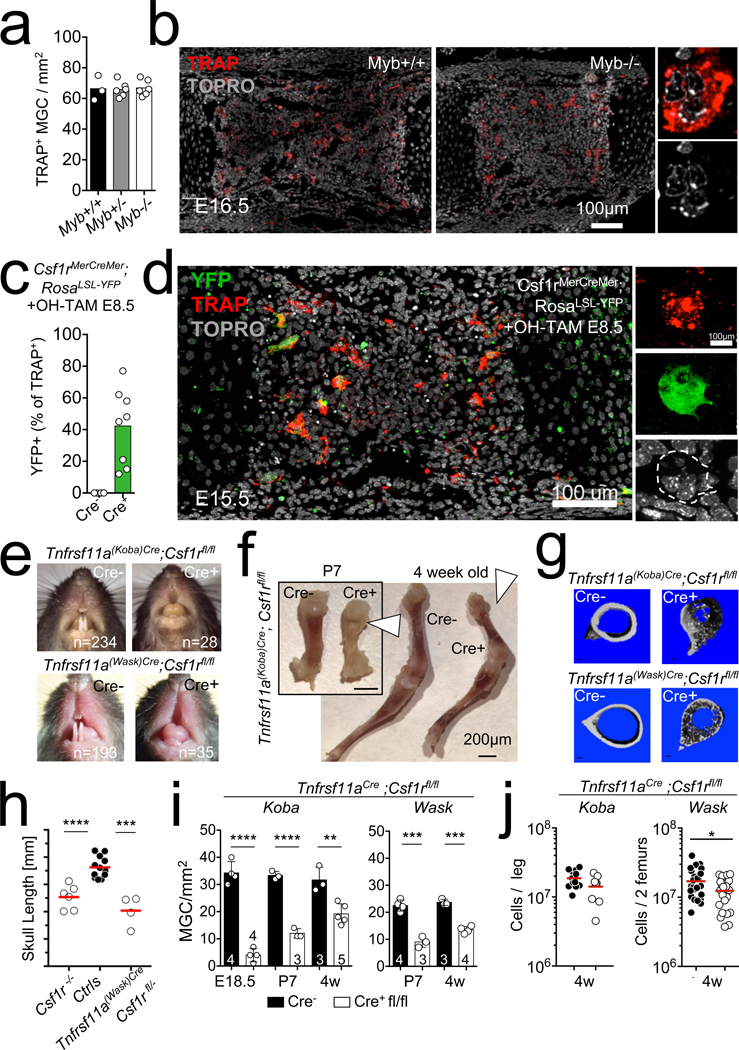
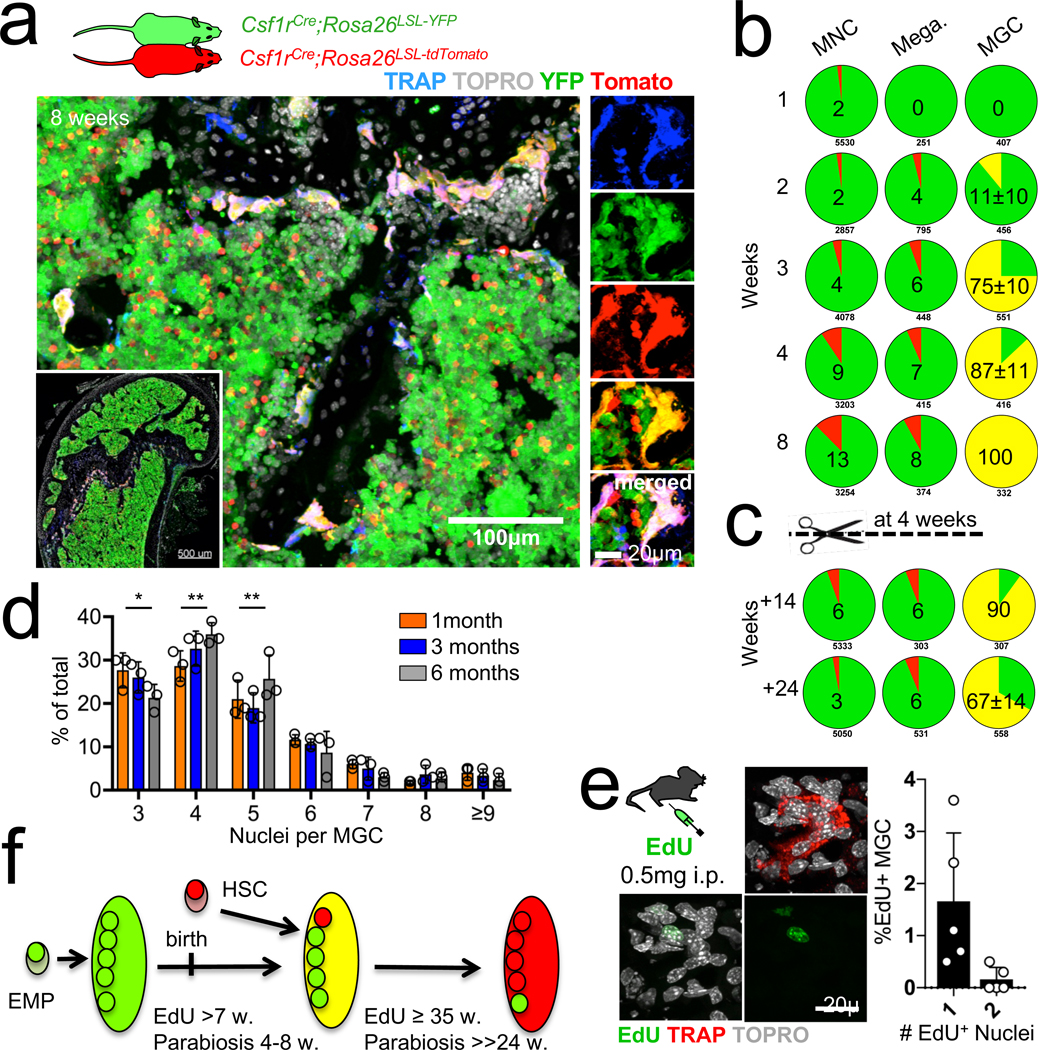
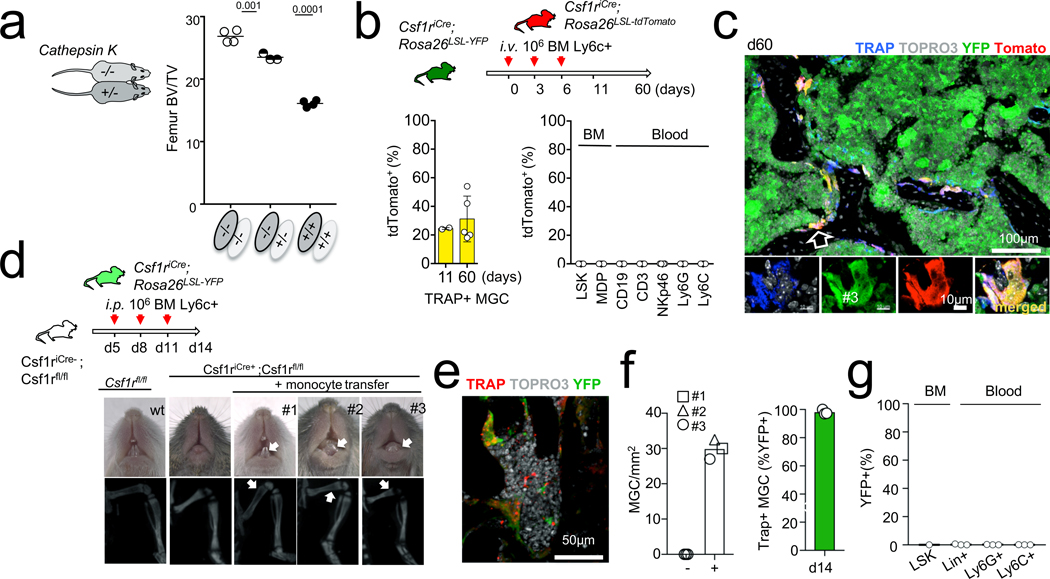
Osteoclasts are multinucleated giant cells that resorb bone, ensuring development and continuous remodelling of the skeleton and the bone marrow haematopoietic niche. Defective osteoclast activity leads to osteopetrosis and bone marrow failure1-9, whereas excess activity can contribute to bone loss and osteoporosis10. Osteopetrosis can be partially treated by bone marrow transplantation in humans and mice11-18, consistent with a haematopoietic origin of osteoclasts13,16,19 and studies that suggest that they develop by fusion of monocytic precursors derived from haematopoietic stem cells in the presence of CSF1 and RANK ligand1,20. However, the developmental origin and lifespan of osteoclasts, and the mechanisms that ensure maintenance of osteoclast function throughout life in vivo remain largely unexplored. Here we report that osteoclasts that colonize fetal ossification centres originate from embryonic erythro-myeloid progenitors21,22. These erythro-myeloid progenitor-derived osteoclasts are required for normal bone development and tooth eruption. Yet, timely transfusion of haematopoietic-stem-cell-derived monocytic cells in newborn mice is sufficient to rescue bone development in early-onset autosomal recessive osteopetrosis. We also found that the postnatal maintenance of osteoclasts, bone mass and the bone marrow cavity involve iterative fusion of circulating blood monocytic cells with long-lived osteoclast syncytia. As a consequence, parabiosis or transfusion of monocytic cells results in long-term gene transfer in osteoclasts in the absence of haematopoietic-stem-cell chimerism, and can rescue an adult-onset osteopetrotic phenotype caused by cathepsin K deficiency23,24. In sum, our results identify the developmental origin of osteoclasts and a mechanism that controls their maintenance in bones after birth. These data suggest strategies to rescue osteoclast deficiency in osteopetrosis and to modulate osteoclast activity in vivo.
Conflict of interest statement
Figures
Similar articles
-
Committed osteoclast precursors colonize the bone and improve the phenotype of a mouse model of autosomal recessive osteopetrosis.J Bone Miner Res. 2010 Jan;25(1):106-13. doi: 10.1359/jbmr.090715. J Bone Miner Res. 2010. PMID: 20091929
-
Hematopoietic Stem Cell-Targeted Neonatal Gene Therapy with a Clinically Applicable Lentiviral Vector Corrects Osteopetrosis in oc/oc Mice.Hum Gene Ther. 2019 Nov;30(11):1395-1404. doi: 10.1089/hum.2019.047. Epub 2019 Jul 3. Hum Gene Ther. 2019. PMID: 31179768
-
The origin of osteoclasts.Immunobiology. 1982 Apr;161(3-4):193-203. doi: 10.1016/S0171-2985(82)80074-0. Immunobiology. 1982. PMID: 7047369 Review.
-
Osteopetrosis in mice lacking haematopoietic transcription factor PU.1.Nature. 1997 Mar 6;386(6620):81-4. doi: 10.1038/386081a0. Nature. 1997. PMID: 9052784
-
Recent developments in the understanding of the pathophysiology of osteopetrosis.Eur J Endocrinol. 1996 Feb;134(2):143-56. doi: 10.1530/eje.0.1340143. Eur J Endocrinol. 1996. PMID: 8630510 Review.
Cited by
-
Oncostatin M: Dual Regulator of the Skeletal and Hematopoietic Systems.Curr Osteoporos Rep. 2024 Feb;22(1):80-95. doi: 10.1007/s11914-023-00837-z. Epub 2024 Jan 10. Curr Osteoporos Rep. 2024. PMID: 38198032 Free PMC article. Review.
-
The serine synthesis pathway drives osteoclast differentiation through epigenetic regulation of NFATc1 expression.Nat Metab. 2024 Jan;6(1):141-152. doi: 10.1038/s42255-023-00948-y. Epub 2024 Jan 10. Nat Metab. 2024. PMID: 38200114 Free PMC article.
-
Immune mediated support of metastasis: Implication for bone invasion.Cancer Commun (Lond). 2024 Sep;44(9):967-991. doi: 10.1002/cac2.12584. Epub 2024 Jul 14. Cancer Commun (Lond). 2024. PMID: 39003618 Free PMC article. Review.
-
Inhibition of Sirtuin 3 prevents titanium particle-induced bone resorption and osteoclastsogenesis via suppressing ERK and JNK signaling.Int J Biol Sci. 2021 Apr 3;17(5):1382-1394. doi: 10.7150/ijbs.53992. eCollection 2021. Int J Biol Sci. 2021. PMID: 33867853 Free PMC article.
-
Cell surface-bound La protein regulates the cell fusion stage of osteoclastogenesis.Nat Commun. 2023 Feb 4;14(1):616. doi: 10.1038/s41467-023-36168-x. Nat Commun. 2023. PMID: 36739273 Free PMC article.
References
-
- Lacey DL et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 93, 165–176 (1998). - PubMed
-
- Yoshida H et al. The murine mutation osteopetrosis is in the coding region of the macrophage colony stimulating factor gene. Nature 345, 442–444 (1990). - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
