

mTORC2-mediated PDHE1α nuclear translocation links EBV-LMP1 reprogrammed glucose metabolism to cancer metastasis in nasopharyngeal carcinoma
- PMID: 30745576
- PMCID: PMC6756087
- DOI: 10.1038/s41388-019-0749-y
mTORC2-mediated PDHE1α nuclear translocation links EBV-LMP1 reprogrammed glucose metabolism to cancer metastasis in nasopharyngeal carcinoma
Abstract
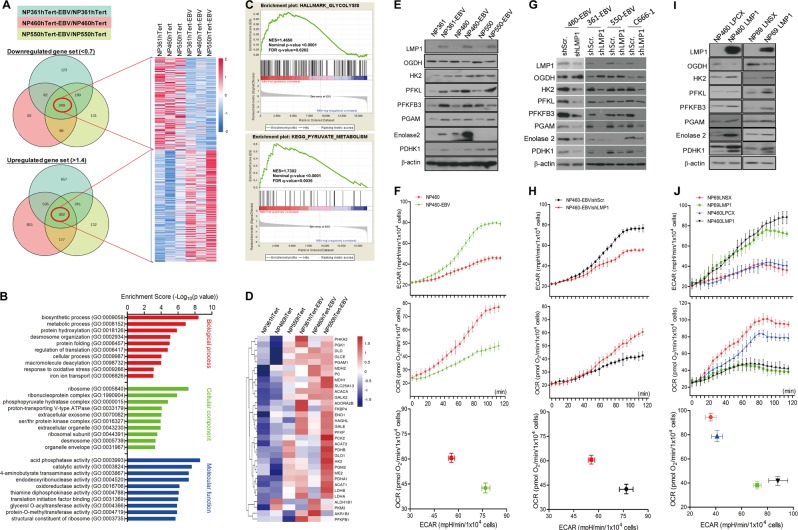
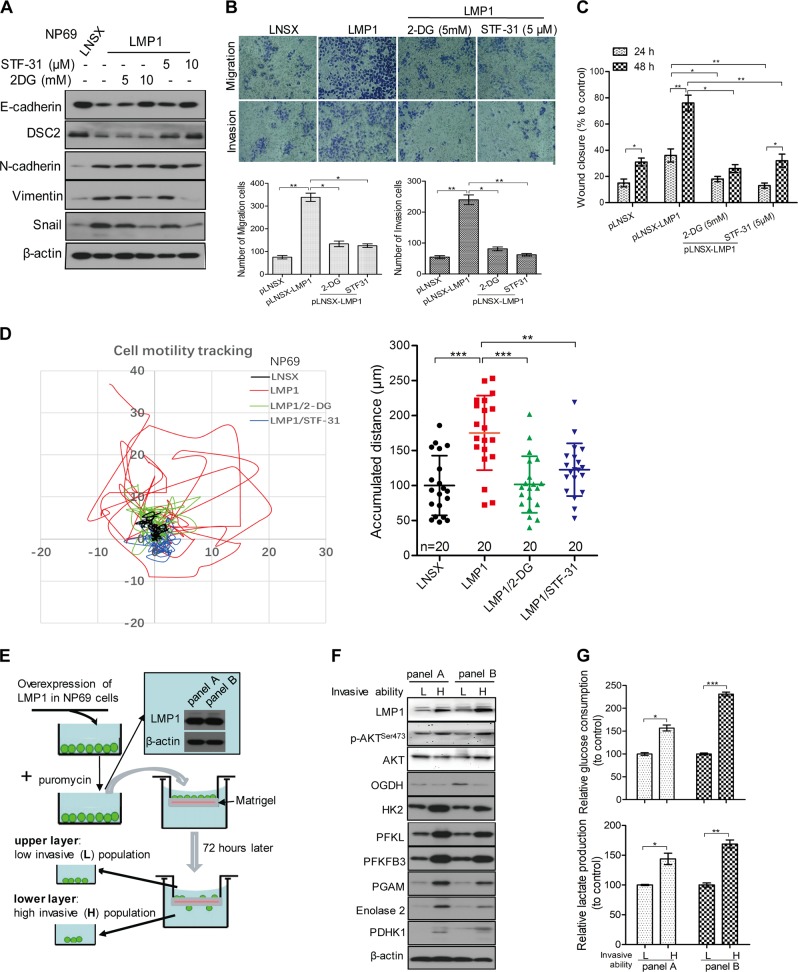
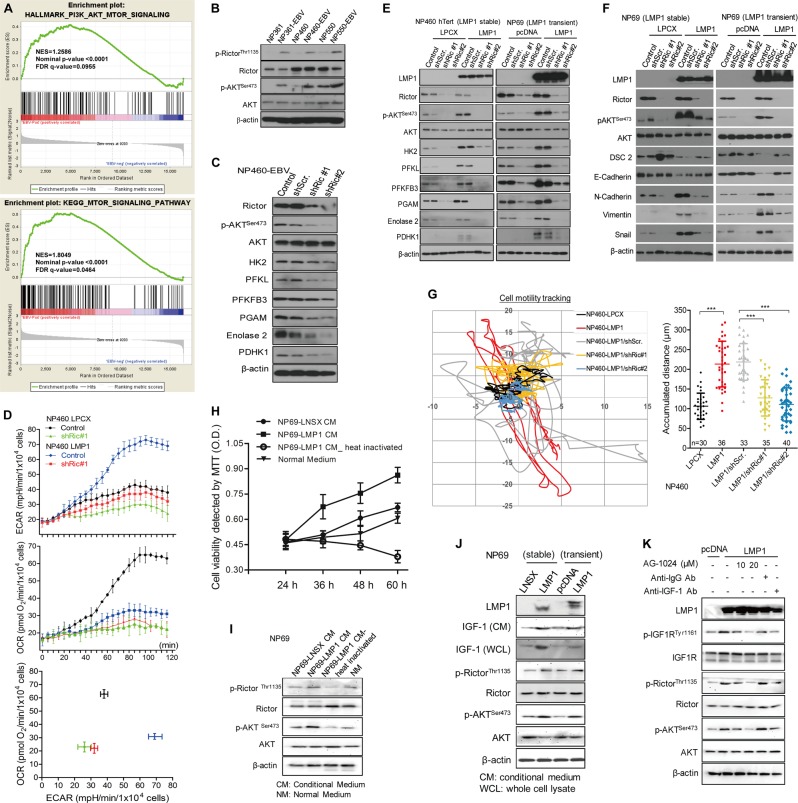
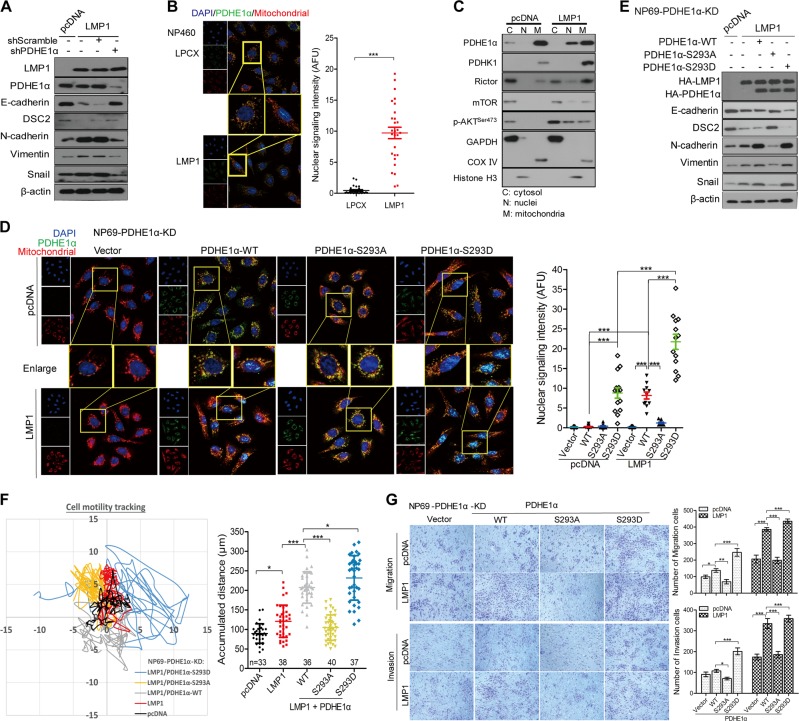
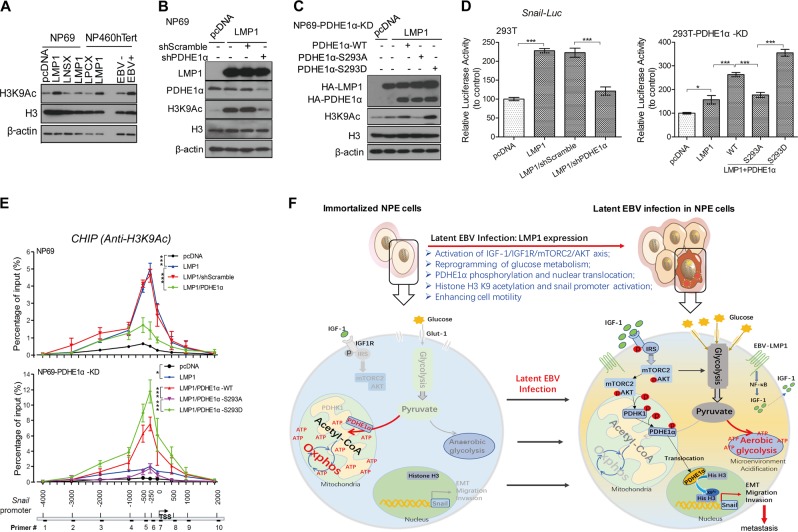
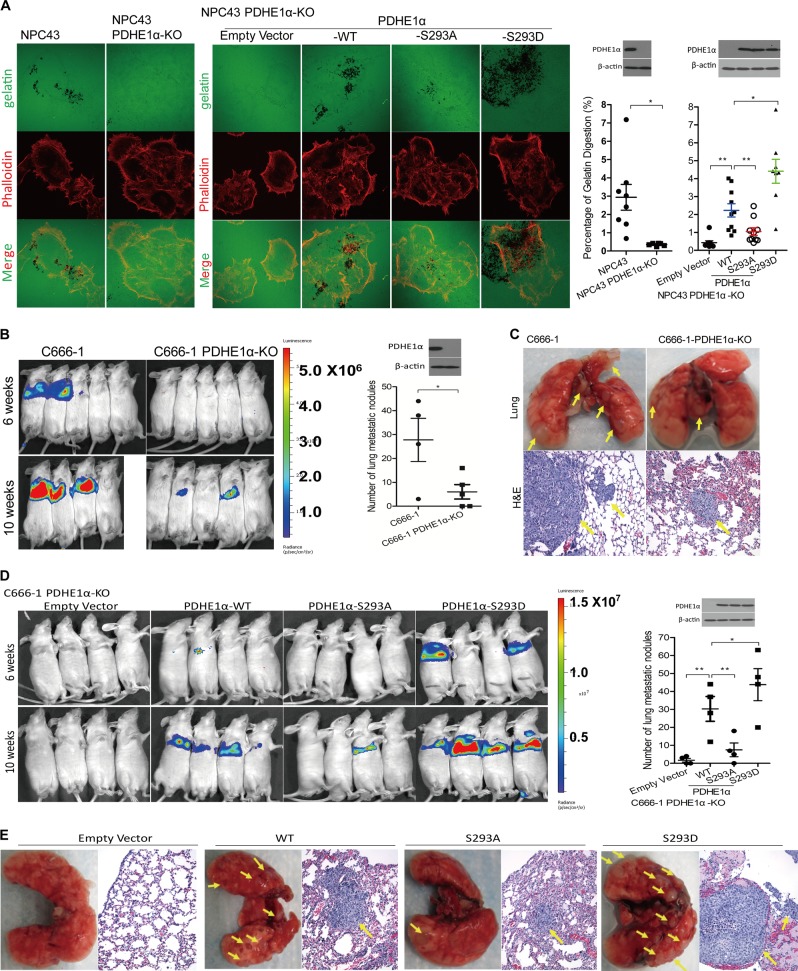
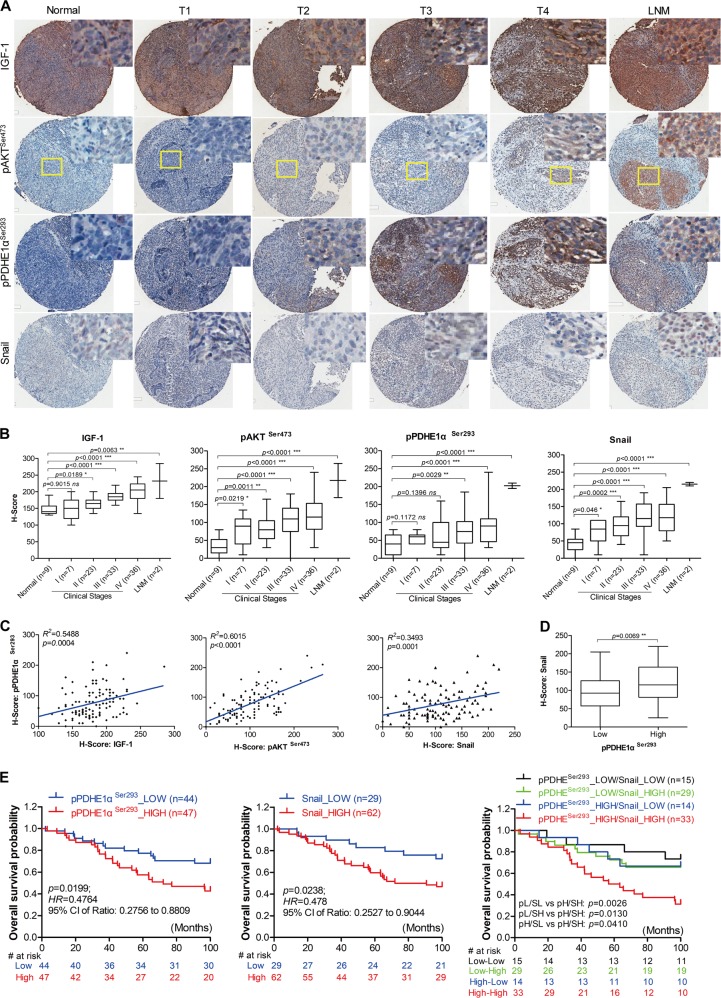
EBV infection of preinvasive nasopharyngeal epithelium is believed to be an initiation step during pathogenesis of nasopharyngeal carcinoma (NPC), but the mechanisms remain poorly understood. Here we report a novel mechanism driving NPC metastasis through the EBV-encoded LMP1-mediated metabolic reprogramming, via activation of IGF1-mTORC2 signaling and nuclear acetylation of the Snail promoter by the PDHE1α, an enzyme involved in glucose metabolism. Mechanistically, EBV-LMP1 increases the cellular secretion of IGF1 which promotes phosphorylation of IGF1R to activate mTORC2/AKT signaling linking glucose metabolism to cell motility. LMP1 expression facilitates translocation of mitochondrial PDHE1α into the nucleus in a phosphorylation-dependent manner at Ser293 residue. Functionally, nuclear PDHE1α promotes H3K9 acetylation on the Snail promoter to enhance cell motility, thereby driving cancer metastasis. Importantly, the IGF1/mTORC2/PDHE1α/Snail axis correlates significantly with disease progression and poor prognosis in NPC patients. This study highlights the functional importance of IGF1-mTORC2-PDHE1α signaling mediated by EBV-LMP1 in NPC pathogenesis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Activation of sterol regulatory element-binding protein 1 (SREBP1)-mediated lipogenesis by the Epstein-Barr virus-encoded latent membrane protein 1 (LMP1) promotes cell proliferation and progression of nasopharyngeal carcinoma.J Pathol. 2018 Oct;246(2):180-190. doi: 10.1002/path.5130. Epub 2018 Aug 22. J Pathol. 2018. PMID: 29968360 Free PMC article.
-
Drp1-dependent remodeling of mitochondrial morphology triggered by EBV-LMP1 increases cisplatin resistance.Signal Transduct Target Ther. 2020 May 20;5(1):56. doi: 10.1038/s41392-020-0151-9. Signal Transduct Target Ther. 2020. PMID: 32433544 Free PMC article.
-
Activation of the FGFR1 signalling pathway by the Epstein-Barr virus-encoded LMP1 promotes aerobic glycolysis and transformation of human nasopharyngeal epithelial cells.J Pathol. 2015 Oct;237(2):238-48. doi: 10.1002/path.4575. Epub 2015 Aug 3. J Pathol. 2015. PMID: 26096068
-
EBV Infection and Glucose Metabolism in Nasopharyngeal Carcinoma.Adv Exp Med Biol. 2017;1018:75-90. doi: 10.1007/978-981-10-5765-6_6. Adv Exp Med Biol. 2017. PMID: 29052133 Review.
-
Novel roles and therapeutic targets of Epstein-Barr virus-encoded latent membrane protein 1-induced oncogenesis in nasopharyngeal carcinoma.Expert Rev Mol Med. 2015 Aug 18;17:e15. doi: 10.1017/erm.2015.13. Expert Rev Mol Med. 2015. PMID: 26282825 Review.
Cited by
-
A Review: PI3K/AKT/mTOR Signaling Pathway and Its Regulated Eukaryotic Translation Initiation Factors May Be a Potential Therapeutic Target in Esophageal Squamous Cell Carcinoma.Front Oncol. 2022 Apr 28;12:817916. doi: 10.3389/fonc.2022.817916. eCollection 2022. Front Oncol. 2022. PMID: 35574327 Free PMC article. Review.
-
EBV Infection and Its Regulated Metabolic Reprogramming in Nasopharyngeal Tumorigenesis.Front Cell Infect Microbiol. 2022 Jul 1;12:935205. doi: 10.3389/fcimb.2022.935205. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35846746 Free PMC article. Review.
-
TGF-β1 is a regulator of the pyruvate dehydrogenase complex in fibroblasts.Sci Rep. 2020 Oct 21;10(1):17914. doi: 10.1038/s41598-020-74919-8. Sci Rep. 2020. PMID: 33087819 Free PMC article.
-
EBV latent membrane protein 1 augments γδ T cell cytotoxicity against nasopharyngeal carcinoma by induction of butyrophilin molecules.Theranostics. 2023 Jan 1;13(2):458-471. doi: 10.7150/thno.78395. eCollection 2023. Theranostics. 2023. PMID: 36632221 Free PMC article.
-
LMP1 Up-regulates Calreticulin to Induce Epithelial-mesenchymal Transition via TGF-β/Smad3/NRP1 Pathway in Nasopharyngeal Carcinoma Cells.J Cancer. 2020 Jan 1;11(5):1257-1269. doi: 10.7150/jca.37415. eCollection 2020. J Cancer. 2020. PMID: 31956372 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
