

CD44/CD44v6 a Reliable Companion in Cancer-Initiating Cell Maintenance and Tumor Progression
- PMID: 30211160
- PMCID: PMC6122270
- DOI: 10.3389/fcell.2018.00097
CD44/CD44v6 a Reliable Companion in Cancer-Initiating Cell Maintenance and Tumor Progression
Abstract
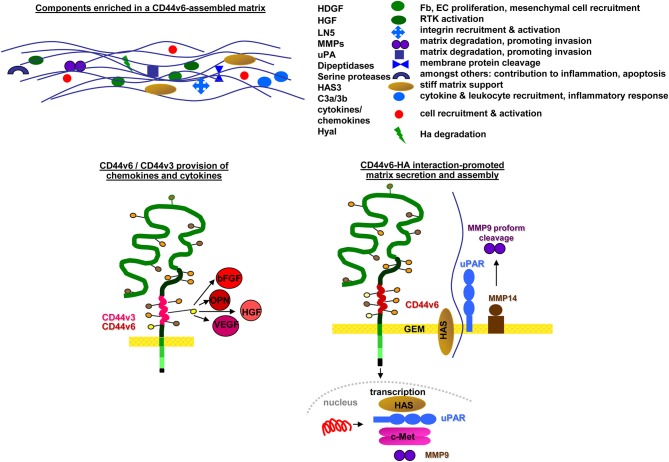
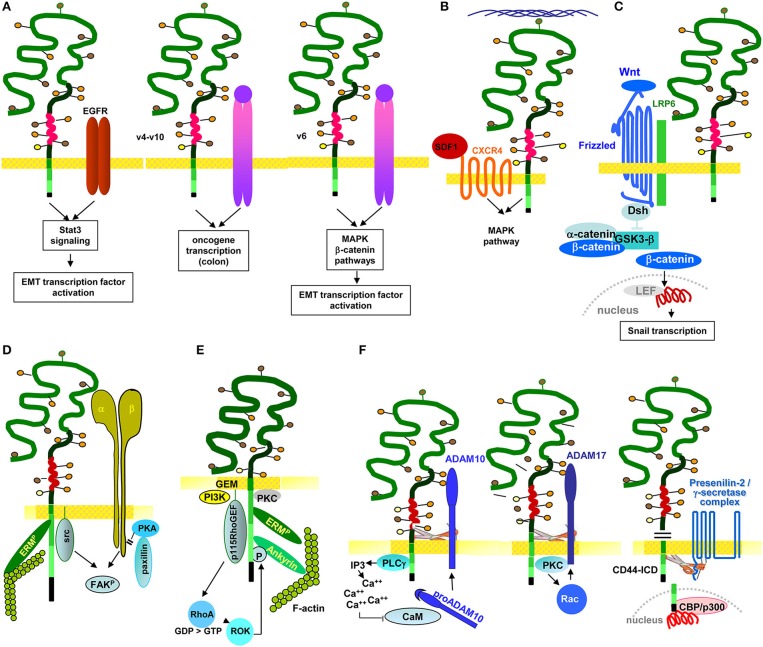
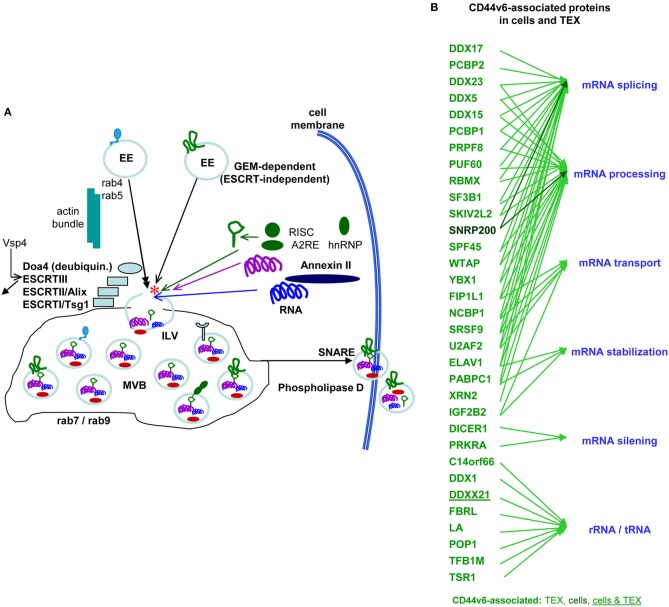
Metastasis is the leading cause of cancer death, tumor progression proceeding through emigration from the primary tumor, gaining access to the circulation, leaving the circulation, settling in distant organs and growing in the foreign environment. The capacity of a tumor to metastasize relies on a small subpopulation of cells in the primary tumor, so called cancer-initiating cells (CIC). CIC are characterized by sets of markers, mostly membrane anchored adhesion molecules, CD44v6 being the most frequently recovered marker. Knockdown and knockout models accompanied by loss of tumor progression despite unaltered primary tumor growth unraveled that these markers are indispensable for CIC. The unexpected contribution of marker molecules to CIC-related activities prompted research on underlying molecular mechanisms. This review outlines the contribution of CD44, particularly CD44v6 to CIC activities. A first focus is given to the impact of CD44/CD44v6 to inherent CIC features, including the crosstalk with the niche, apoptosis-resistance, and epithelial mesenchymal transition. Following the steps of the metastatic cascade, we report on supporting activities of CD44/CD44v6 in migration and invasion. These CD44/CD44v6 activities rely on the association with membrane-integrated and cytosolic signaling molecules and proteases and transcriptional regulation. They are not restricted to, but most pronounced in CIC and are tightly regulated by feedback loops. Finally, we discuss on the engagement of CD44/CD44v6 in exosome biogenesis, loading and delivery. exosomes being the main acteurs in the long-distance crosstalk of CIC with the host. In brief, by supporting the communication with the niche and promoting apoptosis resistance CD44/CD44v6 plays an important role in CIC maintenance. The multifaceted interplay between CD44/CD44v6, signal transducing molecules and proteases facilitates the metastasizing tumor cell journey through the body. By its engagement in exosome biogenesis CD44/CD44v6 contributes to disseminated tumor cell settlement and growth in distant organs. Thus, CD44/CD44v6 likely is the most central CIC biomarker.
Keywords: CD44; EMT; apoptosis resistance; cancer initiating cells; metastasis; migration; tumor exosomes.
Figures


Similar articles
-
CD44v6-competent tumor exosomes promote motility, invasion and cancer-initiating cell marker expression in pancreatic and colorectal cancer cells.Oncotarget. 2016 Aug 23;7(34):55409-55436. doi: 10.18632/oncotarget.10580. Oncotarget. 2016. PMID: 27419629 Free PMC article.
-
Pancreatic cancer-initiating cell exosome message transfer into noncancer-initiating cells: the importance of CD44v6 in reprogramming.J Exp Clin Cancer Res. 2019 Mar 19;38(1):132. doi: 10.1186/s13046-019-1129-8. J Exp Clin Cancer Res. 2019. Retraction in: J Exp Clin Cancer Res. 2025 Feb 28;44(1):78. doi: 10.1186/s13046-025-03339-1. PMID: 30890157 Free PMC article. Retracted.
-
The Pancreatic Cancer-Initiating Cell Marker CD44v6 Affects Transcription, Translation, and Signaling: Consequences for Exosome Composition and Delivery.J Oncol. 2019 Aug 7;2019:3516973. doi: 10.1155/2019/3516973. eCollection 2019. J Oncol. 2019. PMID: 31485223 Free PMC article.
-
CD44 and EpCAM: cancer-initiating cell markers.Curr Mol Med. 2008 Dec;8(8):784-804. doi: 10.2174/156652408786733667. Curr Mol Med. 2008. PMID: 19075676 Review.
-
Exosomes, metastases, and the miracle of cancer stem cell markers.Cancer Metastasis Rev. 2019 Jun;38(1-2):259-295. doi: 10.1007/s10555-019-09793-6. Cancer Metastasis Rev. 2019. PMID: 31030373 Review.
Cited by
-
Immune therapy: a new therapy for acute myeloid leukemia.Blood Sci. 2022 Dec 15;5(1):15-24. doi: 10.1097/BS9.0000000000000140. eCollection 2023 Jan. Blood Sci. 2022. PMID: 36742185 Free PMC article. Review.
-
Association of CD44 gene rs187115 polymorphism with colorectal cancer risk and prognosis in Chinese Han population: a case-control study.Aging (Albany NY). 2019 Nov 3;11(21):9616-9625. doi: 10.18632/aging.102408. Epub 2019 Nov 3. Aging (Albany NY). 2019. PMID: 31682231 Free PMC article.
-
The plasticity of pancreatic cancer stem cells: implications in therapeutic resistance.Cancer Metastasis Rev. 2021 Sep;40(3):691-720. doi: 10.1007/s10555-021-09979-x. Epub 2021 Aug 28. Cancer Metastasis Rev. 2021. PMID: 34453639 Free PMC article. Review.
-
The Role of Proteoglycans in Cancer Metastasis and Circulating Tumor Cell Analysis.Front Cell Dev Biol. 2020 Aug 26;8:749. doi: 10.3389/fcell.2020.00749. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32984308 Free PMC article. Review.
-
Roniciclib down-regulates stemness and inhibits cell growth by inducing nucleolar stress in neuroblastoma.Sci Rep. 2020 Jul 31;10(1):12902. doi: 10.1038/s41598-020-69499-6. Sci Rep. 2020. PMID: 32737364 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous